Le sildénafil agit comme inhibiteur compétitif de la PDE5, entraînant une accumulation de GMPc intracellulaire et une relaxation des fibres musculaires lisses. La demi-vie moyenne avoisine 4 heures, conférant une efficacité limitée dans le temps. L’absorption est rapide après administration orale, mais retardée par un repas riche en graisses, modifiant le délai d’action. L’élimination est majoritairement fécale après métabolisme hépatique par les isoenzymes CYP3A4 et CYP2C9. Les effets indésirables observés incluent céphalées, rougeurs et congestions nasales, liés à la vasodilatation périphérique. Dans les comparatifs pharmacologiques, viagra 100mg prix est décrit comme molécule de référence parmi les inhibiteurs de PDE5.
2106014
Prolonged Methylprednisolone Treatment Suppresses Systemic Inflammation in Patients with Unresolving Acute Respiratory Distress Syndrome Evidence for Inadequate Endogenous Glucocorticoid Secretion and Inflammation-induced Immune Cell Resistance to Glucocorticoids G. Umberto Meduri, Elizabeth A. Tolley, George P. Chrousos, and Frankie Stentz
Memphis Lung Research Program, Divisions of Pulmonary and Critical Care Medicine, Department of Medicine, and Department of Preventive Medicine, University of Tennessee, Memphis, Tennessee; and Pediatric and Reproductive Endocrinology Branch, National Institute of Child Health and Human Development, National Institutes of Health, Bethesda, Maryland
Nuclear factor-B (NF-B) and glucocorticoid receptor-␣ (GR-␣)
that the final outcome is affected by its degree and duration
have diametrically opposed functions in regulating inflammation.
(1). Two cellular signaling pathways are central to the host in-
We investigated whether unresolving acute respiratory distress
flammatory response, the stimulatory nuclear factor-B (NF-B)
syndrome (ARDS) is associated with systemic inflammation-
and the inhibitory glucocorticoid receptor-␣ (GR-␣)-mediated
induced glucocorticoid resistance and whether prolonged methyl- prednisolone administration accelerates the suppression of sys-
NF-B is a heterodimeric protein composed of the DNA-
temic inflammatory indices and normalizes the sensitivity of the
binding proteins p65 and p50 constitutively present in the cy-
immune system to glucocorticoids. Patients enrolled into a ran-
toplasm in an inactive form stabilized by the binding to the in-
domized trial evaluating prolonged methylprednisolone adminis-
hibitory protein IB␣ (4). Cellular activation by a multitude of
tration in unresolving ARDS had serial plasma samples collected
adverse stimuli leads to phosphorylation and proteolytic deg-
before and after randomization. In the plasma, we measured the
radation of IB␣ (4). The liberated NF-B then translocates
concentrations of tumor necrosis factor-␣ (TNF-␣), interleukins (IL)
into the nucleus and binds to promoter regions of target genes
IL-1 and IL-6, adrenocorticotropic hormone (ACTH), and cortisol. The ability of patient plasma to influence the NF-B and GR-signal
to initiate the transcription of multiple cytokines including tu-
transduction systems of normal peripheral blood leukocytes (PBL)
mor necrosis factor-␣ (TNF-␣); the interleukins (IL) IL-1,
was examined. Patients treated with methylprednisolone had pro-
IL-2, IL-6; and chemokines such as IL-8, cell adhesion mole-
gressive and sustained reductions of TNF-␣, IL-1, IL-6, ACTH, and
cules, and inflammation-associated enzymes (4). Products of
cortisol concentrations over time. Normal PBL exposed to plasma
the genes that are stimulated by NF-B activate this transcrip-
samples collected during methylprednisolone exhibited significant
tion factor. Thus, TNF-␣ and IL-1 both activate and are acti-
progressive increases in all aspects of GR-mediated activity and
vated by NF-B by forming a positive regulatory loop that
significant reductions in NF-B DNA-binding and transcription of
amplifies and perpetuates inflammation (5). TNF-␣ and IL-1. These findings provide support for the presence
Glucocorticoid hormones (GC), produced by the adrenal
of endogenous glucocorticoid inadequacy in the control of inflam-
cortices, are the most important physiologic inhibitors of in-
mation and systemic inflammation-induced peripheral glucocorti-
flammation. GC exert most of their effects by activating ubiq-
coid resistance in ARDS. Prolonged methylprednisolone adminis-
uitously distributed cytoplasmic heat shock protein–complexed
tration accelerated the resolution of both systemic inflammation
glucocorticoid receptors (GR) with formation of GC—GR-␣
and peripheral acquired glucocorticoid resistance in ARDS.
complexes (6). It is now appreciated that the GC—GR-␣ com-plexes modulate transcription in a hormone-dependent man-
Keywords: acute respiratory distress syndrome; glucocorticoid recep-
ner by binding to glucocorticoid response elements (GRE) in
tors; IB␣; methylprednisolone; nuclear factor-B
the promoters of glucocorticoid responsive genes and by inter-
Excessive systemic inflammation is the pathophysiologic hall-
fering with the activity of other transcription factors such as
mark of pulmonary and extrapulmonary organ dysfunction in
NF-B on genes regulated by these factors (7). GR-mediated
patients with acute respiratory distress syndrome (ARDS) (1),
transcriptional interference is achieved by five important
a frequent cause of hypoxemic respiratory failure associated
mechanisms: (1) by physically interacting with the p65 subunit
with a 40% to 60% mortality (2). It is now appreciated that
and formation of an inactive (GR␣–NF-B) complex (6); (2)
systemic inflammation in ARDS is sustained over time and
by inducing the transcription of the inhibitory protein IB␣gene (6, 8, 9); (3) by blocking degradation of IB␣ via enhancedsynthesis of IL-10 (10); (4) by impairing TNF-␣Ϫinduced deg-radation of IB␣ (11); and (5) by competing for limited
(Received in original form June 6, 2001; accepted in final form January 22, 2002)
amounts of GR␣ coactivators such as CREB-binding proteinand steroid receptor coactivator-1 (12).
This study was supported by the Assisi Foundation of Memphis and the BaptistMemorial Health Care Foundation.
Endogenous glucocorticoids are not always effective in sup-
Correspondence and requests for reprints should be addressed to Dr. G. Um-
pressing life-threatening systemic inflammation, even though
berto Meduri, University of Tennessee Health Science Center, Division of Pulmo-
the degree of cortisolemia frequently correlates with severity
nary and Critical Care Medicine, 956 Court Avenue, Room H316, Memphis, TN
of illness and mortality rate (13–16). Failure to suppress in-
flammation could be due to inadequacy of, and/or tissue resis-
This article has an online data supplement, which is accessible from this issue’s
tance to, the concentrations and durations of endogenous glu-
table of contents online at www.atsjournals.org
cocorticoid elevations, which allow the systemic inflammatory
Am J Respir Crit Care Med Vol 165. pp 983–991, 2002 DOI: 10.1164/rccm.2106014
response to go awry (17). We have recently reported a signifi-
Internet address: www.atsjournals.org
cant physiologic and survival benefit when prolonged gluco-
AMERICAN JOURNAL OF RESPIRATORY AND CRITICAL CARE MEDICINE
corticoid treatment at moderate doses was administered late
TABLE 1. CLINICAL VARIABLES AT THE ONSET OF ARDS
(9 Ϯ 3 d) in the course of ARDS to patients failing to improve
(18). We hypothesized that if endogenous glucocorticoid inad-equacy and/or peripheral tissue resistance are important patho-
physiologic factors in a dysregulated, protracted systemic in-
flammatory response in ARDS, then prolonged glucocorticoid
therapy may be useful, not as an antiinflammatory treatment
per se, but as hormonal supplementation necessary to compen-
sate for the host’s inability to produce appropriately elevated
amounts of cortisol for the degree of peripheral glucocorticoid
No significant difference was found between the two groups for any variable.
In this study, we tested the hypothesis that prolonged me-
* APACHE III score at intensive care unit admission. †
thylprednisolone versus placebo administration in patients with
unresolving ARDS suppresses inflammation and/or correctsthe glucocorticoid resistance of the inflammatory response of
Plasma Collection and Description of the Ex Vivo
these patients. To test this hypothesis, we measured serially
Model of ARDS
the plasma levels of TNF-␣, IL-1, IL-6, adrenocorticotropic
Blood samples were obtained on Days 1, 3, 5, 7, and 10 of ARDS, on the
hormone (ACTH), and cortisol in patients with unresolving
day of randomization (Day 0), and on Days 3 (ϩ3), 5 (ϩ5), 7 (ϩ7), and
ARDS treated with methylprednisolone or placebo and ex-
10 (ϩ10) of treatment. All blood samples were obtained from a central
posed a healthy volunteer’s peripheral blood leukocytes (PBL)
venous line or an antecubital venipuncture, collected in a vacutainer
to plasma samples from the patients. In the exposed cells, we
tube containing ethylenediaminetetraacetic acid, placed immediately on
measured upstream and downstream events associated with
ice after collection, and transported to the laboratory for immediate pro-
the NF-B and glucocorticoid transduction cascades as they
cessing. A complete blood cell count with differential was determined
using the Coulter ACT diff Hematology Analyzer (Beckman-Coulter,Miami, FL). Blood samples were centrifuged at 500 ϫ g for 10 minutes
in a refrigerated centrifuge, and plasma was aspirated and aliquoted inplastic storage tubes. All samples were stored at Ϫ80Њ C until assay. Patient Selection
The ex vivo model of ARDS consisted of exposing PBL obtained
from a single healthy volunteer to plasma samples obtained from pa-
The original study was conducted between October 1994 and Novem-
tients with ARDS. Using this ex vivo model, we attempted to simulate
ber 1996 in the intensive care units of Baptist Memorial Medical Cen-
the in vivo milieu, but we realize that there are limitations to the infer-
ter and East hospitals, the Regional Medical Center, and the Univer-
ences that can be made by using this approach.
sity of Tennessee Bowld Medical Center, all in Memphis, TN. Thestudy protocol was approved by each institutional review board, and
Laboratory Methodology
informed consent was obtained before enrollment. An active effortwas made to identify and recruit eligible patients with ARDS. Patients
The laboratory methodology section is available in the online data
at least 18 years old were eligible if they met previously described
supplement and includes the following: isolation of PBL and exposure
ARDS consensus criteria (20) and had a lung injury score (LIS) (21)
to patients’ plasma; determination of plasma cytokine, ACTH, and
у 2.5. The ventilator management followed recently developed guide-
cortisol concentrations; cellular fractionation and protein determina-
lines aimed at limiting plateau pressure to less than 35 cm H
tion; electrophoretic mobility gel shift assay (EMSA); detection of
Positive end-expiratory pressure was increased by 3–5 cm H
NF-B and GR-␣ binding to their response elements; quantification
of EMSA, determination of NF-B subunits, GR-␣, and IB␣ by
2O) to achieve the best lung compliance and oxygen saturation
Western blotting; and quantitative reverse transcription-polymerase
IO less than or equal to 0.6. The presence or absence
of improvement in lung function (as defined by LIS [21]) by Day 7 of
ARDS was used to categorize patients as improvers or nonimproversand ARDS as resolving or unresolving. On mechanical ventilation Day
Statistical Analysis
9 Ϯ 3, 24 nonimprovers were enrolled into a prospective, randomized,
This study has an unbalanced nested factorial design, with groups
double-blind, placebo-controlled trial evaluating prolonged methyl-
(methylprednisolone and placebo) as the main plot effect; patients
prednisolone treatment (18). Randomization was done on a 2:1 basis;
were nested within groups; and time was cross-classified with the
16 patients received methylprednisolone, and 8 received placebo. Data
groups and patients. We made the assumption that the variances of
from 17 of the 24 randomized patients are reported in the present arti-
continuous dependent variables on the assessment days were equal. If
cle. Serial blood samples were available for analysis in 17 patients, 6 of
the variances were not equal or the distributions were positively
whom received placebo and served as control subjects. In 7 of the 24
skewed, data were transformed using natural logarithms. Compari-
randomized patients, blood samples were either not obtained (three
sons were made both within and across groups. For comparisons
patients) or were inadequate for serial measurements (four patients).
within groups, the variance for each dependent variable was esti-
Methylprednisolone or placebo was given daily as intravenous push
mated by the pooled within-patient variance from data measured re-
every six hours (one-fourth of the daily dose) and changed to a single
peatedly over time. For comparisons across groups, that variance was
oral dose when oral intake was restored. If the patient was able to tol-
estimated by the weighted average of the residual mean squares and
erate oral intake and had no obvious gastrointestinal dysfunction (i.e.,
the between-patient mean squares from repeated measures analysis of
diarrhea, etc.), we presumed that the gastrointestinal tract was func-
variance (25). Although constituents in plasma serially collected from
tional. A loading dose of 2 mg/kg was followed by 2 mg/kg/day from
patients were moderately and positively correlated, we assume that
Day 1 to 14, 1 mg/kg/day from Day 15 to 21, 0.5 mg/kg/day from Day
results from cell cultures were independent. For each group, partial
22 to 28, 0.25 mg/kg/day on Days 29 and 30, and 0.125 mg/kg/day on
correlation coefficients among selected intracellular markers, ad-
Days 31 and 32. During the study, components of the LIS (21), and
justed for repeated measurements on patients, were estimated. The
multiple organ dysfunction syndrome score (23) were collected, and
assumptions necessary for this analysis are that (1) the markers are
results are published elsewhere (24).
linearly associated over time and (2) the residuals are independently
Table 1 shows clinical characteristics at the onset of ARDS. The 11
and identically distributed normally with variance 2. For all pre-
patients randomized to methylprednisolone improved LIS by study
planned or a priori contrasts stipulated in the main hypotheses, a sig-
Day 10 and survived. In the placebo group, two patients improved LIS
nificance level of 0.05 was chosen for statistical significance. All hy-
by study Day 10 and survived, whereas four failed to improve LIS. Two
pothesis tests are two-tailed. Data were analyzed using the SAS
of the four nonimprovers died within seven days of randomization.
statistical software package (SAS Institute, Inc., Cary, NC).
Meduri, Tolley, Chrousos, et al.: Methylprednisolone Suppresses Inflammation in Unresolving ARDS
and up to randomization, plasma ACTH and cortisol levelswere similar in both groups. After randomization, the methyl-
Cytokine Concentrations from Plasma of Randomized Patients
prednisolone group had significant and sustained (on Days 5 to
A total of 88 blood specimens was available for analysis, 58
7, p Ͻ 0.005 for both measurements) reductions in plasma
from patients in the methylprednisolone group and 30 from
ACTH and cortisol concentrations, whereas no reductions
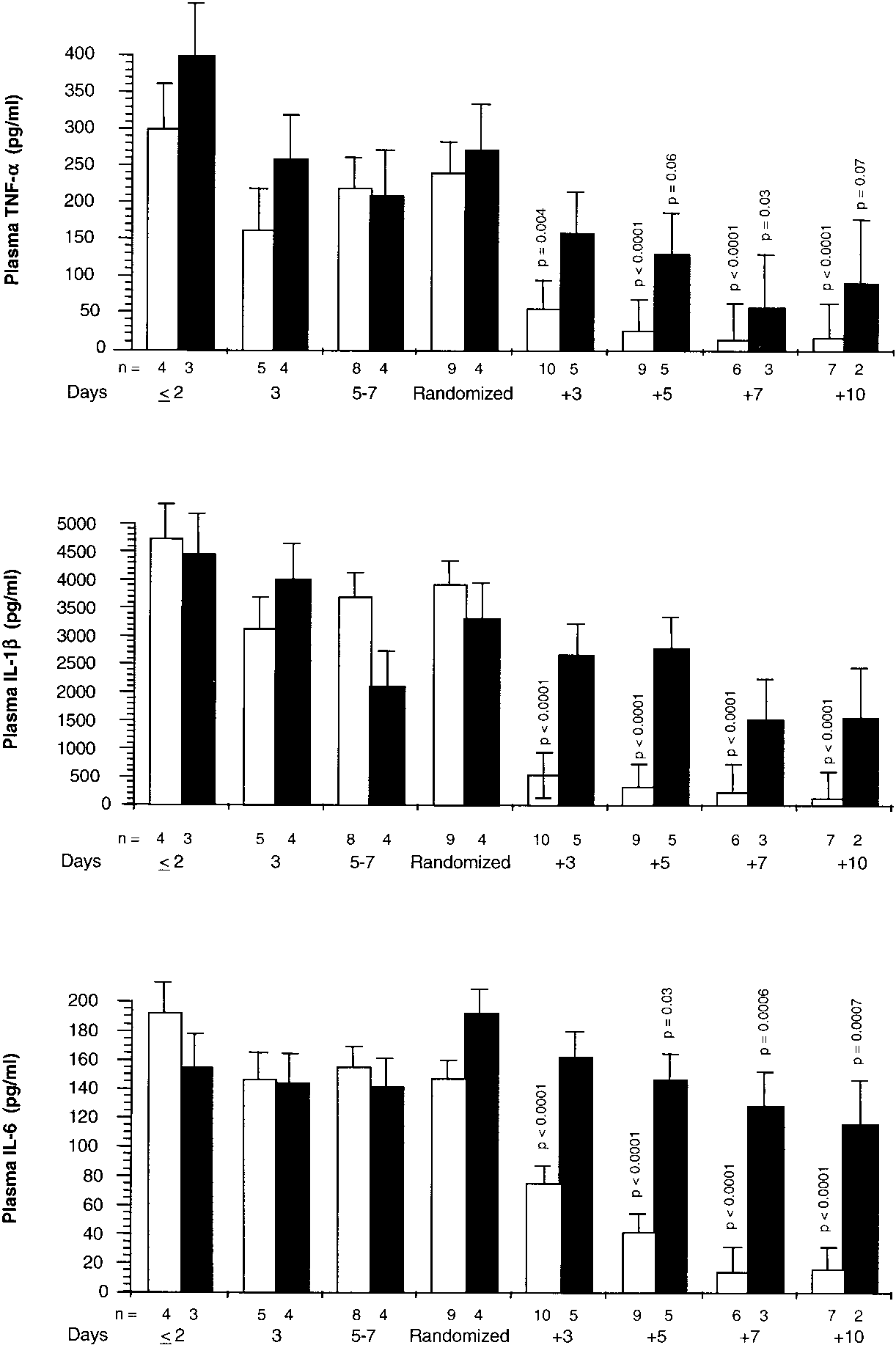
patients in the placebo group. Figure 1 shows the patients’
plasma TNF-␣, IL-1, and IL-6 levels before and after ran-domization. On Day 1 of ARDS and up to randomization,
NF-B and Its Subunits from Cells of a Healthy Volunteer
plasma TNF-␣, IL-1, and IL-6 levels were similar in both
Figure 3 shows NF-B, p65 subunit, and p50 subunit binding
groups. After randomization, plasma TNF-␣, IL-1, and IL-6
to response elements before and after randomization. On Day
levels declined rapidly in the methylprednisolone group (on
1 of ARDS, densities of NF-B, p65 subunit, and p50 subunit
Days 5 to 7, p Ͻ 0.0001 for all three cytokines), whereas the
were similar in cells exposed to plasma from both groups.
control group had lesser reductions in plasma TNF-␣ and IL-6
From Day 1 of ARDS to randomization, densities of all three
and no reduction in plasma IL-1. After randomization, the
proteins increased significantly and similarly in cells exposed
methylprednisolone group had significantly (p Ͻ 0.0001) lower
to plasma from both groups. After randomization, cells
plasma IL-1 concentrations than the control group for each
treated with plasma from the methylprednisolone group ex-
hibited significant progressive reductions in NF-B and itssubunits (on Day 3, p Ͻ 0.02 for all three measurements),
ACTH and Cortisol Concentrations from Plasma of
whereas no changes were observed in those exposed to plasma
Randomized Patients
from the control group. After randomization, significant dif-
Figure 2 shows plasma ACTH and cortisol concentrations in
ferences (p Ͻ 0.01) between the two groups were observed for
patients before and after randomization. On Day 1 of ARDS
Figure 1. Plasma TNF-␣, IL-1, and IL-6 concentrations be- fore and after randomization. Plasma TNF-␣ (top), IL-1 (middle), and IL-6 (bottom) concentrations (mean Ϯ SE) be- fore and after randomization in the methylprednisolone group (open bar) and placebo (closed bar). p Values are taken from analyses of natural logarithms of cytokine values and reflect significances preplanned contrasts of each mean to the mean of the respective group observed on the day of randomization.
AMERICAN JOURNAL OF RESPIRATORY AND CRITICAL CARE MEDICINE
Figure 2. Plasma ACTH and cortisol levels before and after randomization. Plasma ACTH (top) and cortisol (bottom) levels (mean Ϯ SE) before and after randomization in the methylprednisolone group (open bar) and placebo (closed bar). Data were not transformed before statistical analysis. Before randomization, p values reflect significances of pre- planned contrasts of each mean to the mean of the respec- tive group on Day 1 of ARDS. After randomization, p values reflect significances of preplanned contrasts of each mean to the mean of the respective group observed on the day of randomization. GR-␣–Mediated Activity
ization, TNF-␣, IL-1, and IL-10 mRNA levels were similar in
Figure 4 shows cytoplasmic GR-␣ bound and unbound to NF-B
media of cells exposed to plasma from both groups. TNF-␣
before and after randomization. On Day 1 of ARDS, cells ex-
and IL-1 mRNA were significantly lower after exposure to
posed to plasma of both groups had similar densities of GR-␣.
plasma from the methylprednisolone group after randomiza-
After exposure to plasma obtained from Day 1 of ARDS up
tion (p Ͻ 0.0001 for both types of mRNA on each recording),
to randomization, the amounts of GR-␣ bound to NF-B did
whereas cells exposed to plasma from the control group after
not change, whereas the amounts of GR-␣ unbound decreased
randomization had no reductions in TNF-␣ and IL-1 mRNA
significantly. After randomization, exposure to plasma from
levels. Cells exposed to plasma from the methylprednisolone
the methylprednisolone group was associated with significant
group after randomization had significant (p Ͻ 0.0001) and
increases (on Day 3, p Ͻ 0.0001 for both measurements) in
progressive increases in IL-10 mRNA levels, whereas no
GR-␣ bound and unbound, whereas lesser increases were ob-
changes were observed in cells exposed to plasma from the
served in cells exposed to plasma from the control group. Af-
placebo group after randomization until Day 10. After ran-
ter randomization, a significant difference (p Ͻ 0.0001) be-
domization, significant differences (p Ͻ 0.01) between the two
tween the two groups was observed for cytoplasmic GR-␣
groups were observed for IL-1 and IL-10 mRNA levels for
bound and unbound to NF-B for each recorded interval.
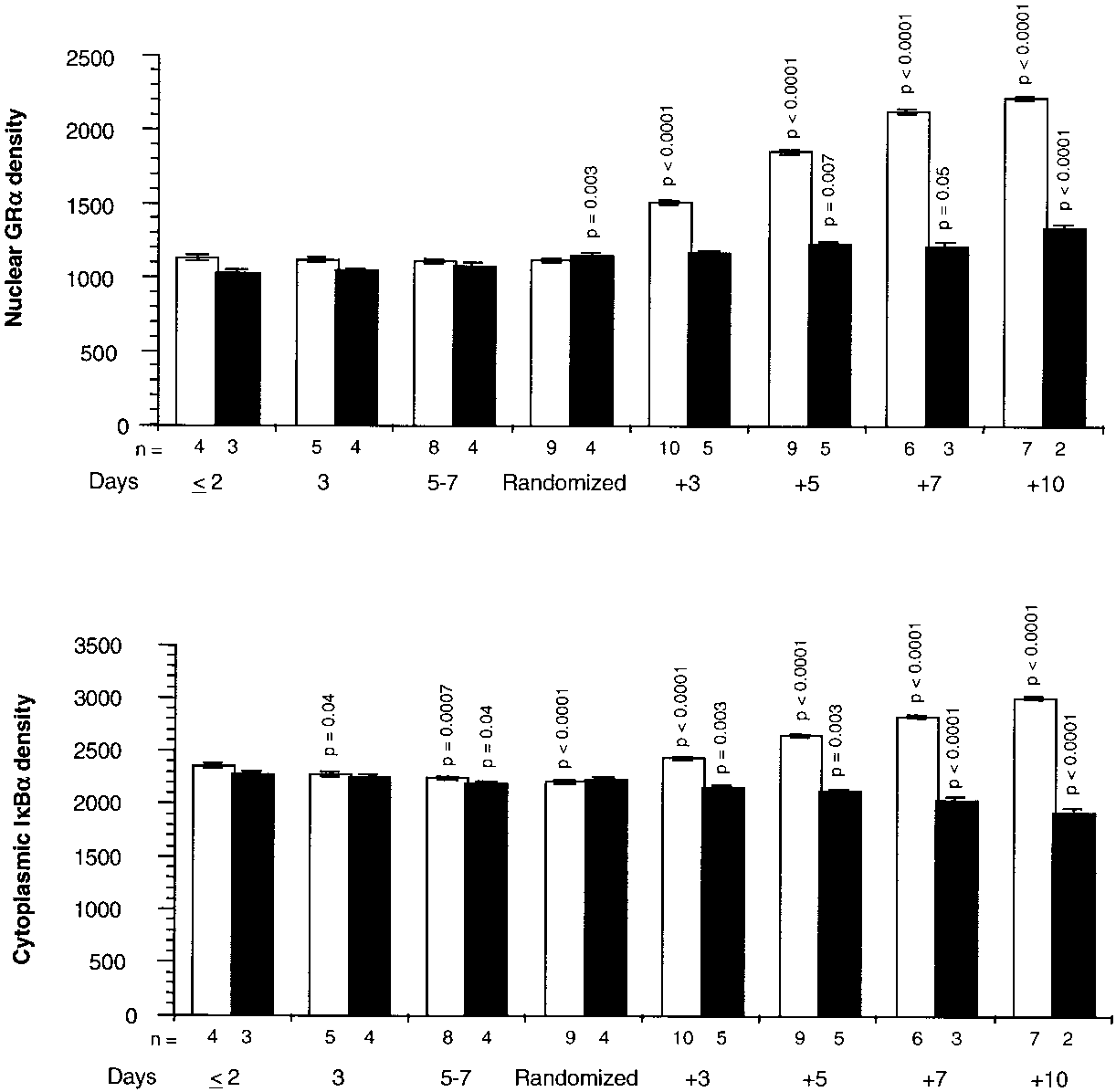
Figure 5 shows cytoplasmic IB␣ and GR-␣ binding to re-
sponse elements. On Day 1 of ARDS and up to randomiza-
Relations among Selected Variables after Randomization
tion, cells exposed to plasma from both groups had similar
Figure 7 depicts the relations on natural logarithmic scales be-
densities of cytoplasmic IB␣ and GR-␣ bound to the re-
tween mean levels of nuclear NF-B and nuclear GR-␣ (top)
sponse elements. After randomization, exposure to plasma
and between mean levels of nuclear NF-B and cytoplasmic
from the methylprednisolone group was associated with sig-
GR-␣ bound to NF-B (bottom), a factor affecting the translo-
nificant increases (on Day 3, p Ͻ 0.0001 for both measure-
cation of activated NF-B to the nucleus. Untransformed
ments) in cytoplasmic IB␣ and GR-␣ bound to the response
means of these transcription factors were depicted separately
elements. In contrast, cells exposed to plasma from the control
in Figures 3 and 4. After natural logarithmic transformation
group had lesser increases in GR␣ bound to the response ele-
and adjustment for repeated measurements, partial correla-
ments and significant reductions in cytoplasmic IB␣ densi-
tions among responses to plasma from the methylprednisolone
ties. After randomization, a significant difference (p Ͻ 0.0001)
group were Ϫ0.92 (p Ͻ 0.0001) both for nuclear NF-B and
between the two groups was observed for cytoplasmic IB␣
nuclear GR-␣ and for nuclear NF-B and cytoplasmic GR-␣
and GR-␣ binding to response elements for each recorded in-
bound to NF-B. For responses to plasma from the placebo
group, no significant relationship was found between nuclearNF-B and nuclear GR-␣ (r ϭ 0.11; p ϭ 0.70) or between NF-B
Cytokine Transcription
and cytoplasmic GR-␣ bound to NF-B (r ϭ 0.33; p ϭ 0.23).
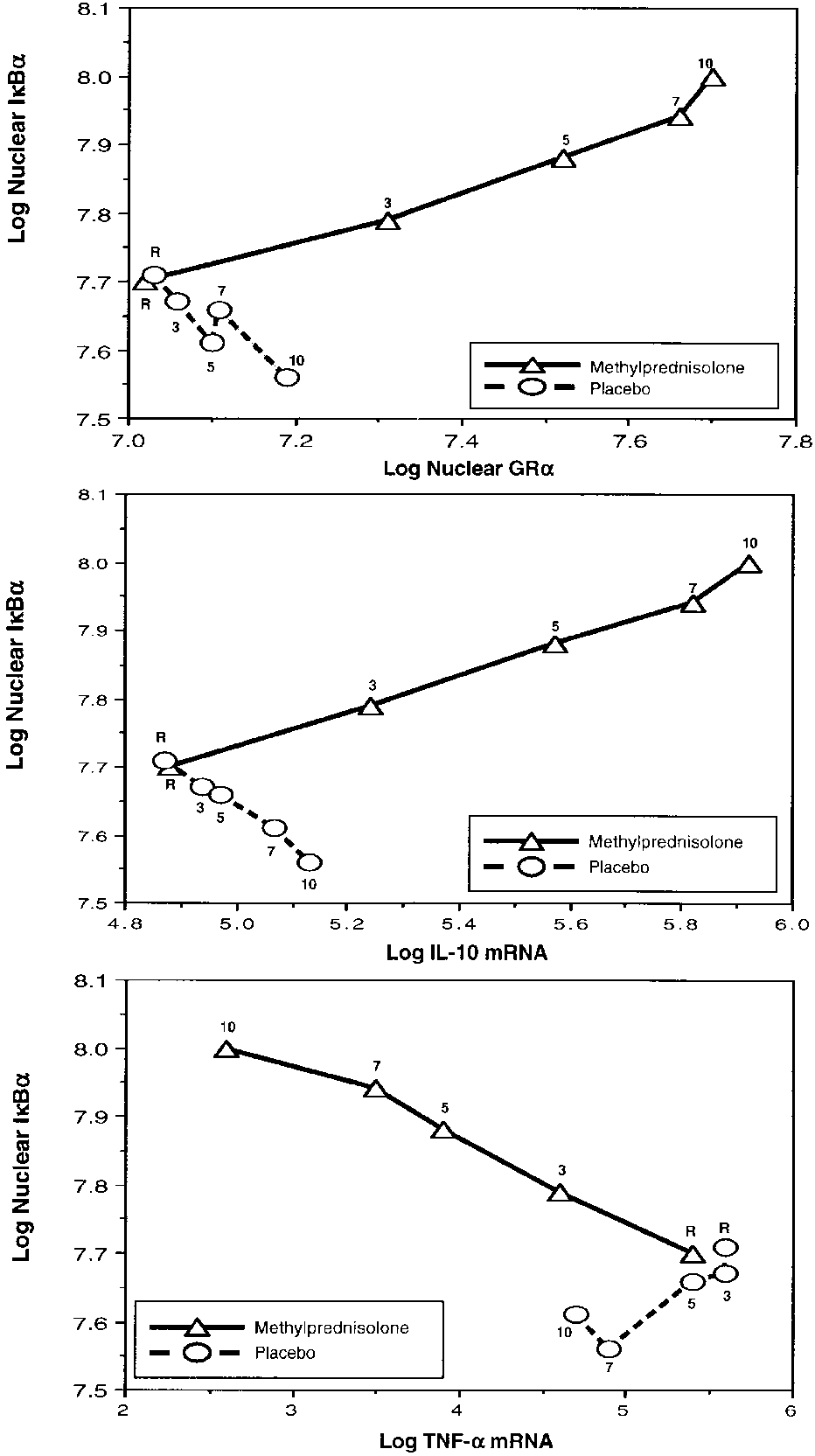
Figure 6 shows TNF-␣, IL-1, and IL-10 mRNA levels mea-
Figure 8 depicts the relations on natural logarithmic scales
sured in media of cells exposed to plasma obtained before and
between mean levels of IB␣ and factors affecting its forma-
after randomization. On Day 1 of ARDS and up to random-
tion (nuclear GR-␣) and degradation (IL-10 mRNA and TNF-␣
Meduri, Tolley, Chrousos, et al.: Methylprednisolone Suppresses Inflammation in Unresolving ARDS
Figure 3. NF-B, p65 subunit, and p50 subunit binding to
B response elements before and after randomization. In-tracellular changes observed by exposing PBL of a healthyvolunteer to plasma samples collected before and after ran-domization. Values are expressed as mean Ϯ SE. NF-B (top),p65 subunit (middle), and p50 subunit (bottom) densities inthe methylprednisolone group (open bar) and placebo (closedbar). NF-B nuclear levels in the PBL were determined byEMSA using chemiluminescent detection. NF-B p65 andp50 subunits were determined by Western blot. Values forNF-B and for the two subunits binding were transformedto natural logarithms before analysis, and p values weretaken from these analyses. Before randomization, p valuesreflect significance of preplanned contrasts of each meanto the mean of the respective group on Day 1 of ARDS. Af-ter randomization, p values reflect significances of pre-planned contrasts of each mean to the mean of the respec-tive group observed on the day of randomization.
mRNA). Untransformed means of these factors were depicted
lected during methylprednisolone versus placebo treatment
separately in Figures 5 and 6. After natural logarithmic trans-
also exhibited rapid, progressive significant increases in GR-
formation and adjustment for repeated measurements, partial
␣–mediated activities (GR-␣ binding to NF-B, GR-␣ binding
correlations were ϩ0.97 (p Ͻ 0.0001) between IB␣ and nu-
to GRE DNA, stimulation of inhibitory protein IB␣, and
clear GR-␣, ϩ0.98 (p Ͻ 0.0001) between IB␣ and IL-10
stimulation of IL-10 transcription) and significant reductions
mRNA, and Ϫ0.95 (p Ͻ 0.0001) between IB␣ and TNF-␣
in NF-B binding and transcription of TNF-␣ and IL-1.
mRNA. In contrast, for responses to plasma from the placebo
These findings provide strong mechanistic evidence for the
group, the partial correlation coefficients were Ϫ0.73 (p ϭ
efficacy of prolonged methylprednisolone treatment in unre-
0.003) between IB␣ and nuclear GR-␣, Ϫ0.85 (p Ͻ 0.0001)
between IB␣ and IL-10 mRNA, and ϩ0.27 (p ϭ 0.33) be-
In the observation period before randomization, the bio-
tween IB␣ and TNF-␣ mRNA. Figure E1 in the web reposi-
logic and physiologic characteristics of the methylpredniso-
tory shows the EMSA of nuclear extract of NF-B over time
lone and placebo groups were similar. Patients had persistent
from one patient randomized to methylprednisolone and from
elevations in plasma concentrations of inflammatory (TNF-␣,
IL-1, and IL-6) cytokines and hypothalamic-pituitary-adre-nal (HPA)-axis (ACTH and cortisol) hormones and similar se-verity of organ dysfunction scores. We hypothesized that inade-
DISCUSSION
quate secretion of cortisol and/or immune tissue resistance to
Patients treated with methylprednisolone had rapid, progres-
endogenous glucocorticoids might explain the observed fail-
sive, and sustained reductions in plasma TNF-␣, IL-1, IL-6,
ure to suppress inflammation in the presence of persistently
ACTH, and cortisol concentrations over time. These were as-
elevated ACTH and cortisol concentrations. Because the GR
sociated with parallel improvements in pulmonary and extra-
ultimately controls GC-mediated activity, anything that af-
pulmonary organ dysfunction scores (previously reported in
fects its binding affinity, concentration, transport to the nu-
reference [18]). Normal PBL exposed to plasma samples col-
cleus, interactions with GRE, or other relevant transcription
AMERICAN JOURNAL OF RESPIRATORY AND CRITICAL CARE MEDICINE
Figure 4. Cytoplasmic GR bound and unbound to NF-B before and after randomization. Intracellular changes ob- served by exposing PBL of a healthy volunteer to plasma samples collected before and after randomization. Values are expressed as mean Ϯ SE. Cytoplasmic GR-␣ bound (top) and unbound (bottom) to NF-B densities in the methyl- prednisolone group (open bar) and placebo (closed bar). Cy- toplasmic GR-␣ bound and unbound to NF-B were deter- mined by Western blot using antigen specific antibodies and fluorescent detection. Bound values were transformed to natural logarithms before analysis, and p values were taken from this analysis. Before randomization, p values re- flect significances of preplanned contrasts of each mean to the mean of the respective group on Day 1 of ARDS. After randomization, p values reflect significance of preplanned contrasts of each mean to the mean of the respective group on the day of randomization.
factors and coregulators can ultimately affect the response of
Recent in vitro studies have shown that cytokines may in-
cells to glucocorticoids (6). GR-mediated resistance was origi-
duce resistance to glucocorticoids by reducing GR-␣ binding
nally described as a primary inherited familial syndrome (26,
affinity to cortisol and/or GRE (34–36). Such abnormalities of
27) and was recently recognized as an acquired condition.
GR-␣ function were demonstrated in T cells incubated with a
Among others, acquired immune tissue-specific GR resistance
combination of IL-2 and IL-4 (35), IL-1, IL-6, and interferon-␥
has been described in patients with asthma (28–31), acquired
(34), or IL-13 (36). Glucocorticoid resistance was induced in a
immunodeficiency syndrome (32), and severe sepsis (33).
cytokine concentration-dependent fashion and was reversed
Figure 5. GR binding to GRE and densities of cytoplasmic IB␣ before and after randomization. Intracellular changes observed by exposing PBL of a healthy volunteer to plasma samples collected before and after randomization. Values are expressed as mean Ϯ SE. GR-␣ binding to response ele- ments (top) and levels of cytoplasmic IB␣ (bottom) in the methylprednisolone (open bar) and placebo (closed bar) groups. Densities of GR-␣ binding to response elements were determined by EMSA, and levels of cytoplasmic IB␣ were determined by Western blot. Data were not trans- formed before statistical analysis. Before randomization, p values reflect significance of preplanned contrasts of each mean to the mean of the respective group on Day 1 of ARDS. After randomization, p values reflect significances of preplanned contrasts of each mean to the mean of the re- spective group on the day of randomization.
Meduri, Tolley, Chrousos, et al.: Methylprednisolone Suppresses Inflammation in Unresolving ARDS
Figure 6. Messenger RNA TNF-␣, IL-1, and IL-10 before and after randomization. Intracellular changes observed by exposing PBL of a healthy volunteer to plasma samples col- lected before and after randomization. Values are expressed as mean Ϯ SE. Messenger RNA TNF-␣ (top), IL-1 (middle), and IL-10 (bottom) levels in the methylprednisolone (open bar) and placebo (closed bar) groups. Values for mRNA of TNF-␣, IL-1, and IL-1 were transformed to natural loga- rithms before analysis, and p values were taken from this analysis. Before randomization, p values reflect significances of preplanned contrasts of each mean to the mean of the respective group on Day 1 of ARDS. After randomization, the p values reflect significances of preplanned contrasts of each mean to the mean of the respective group on the day of randomization.
by the removal of cytokines (35). GR-mediated resistance in
31), and our findings support this hypothesis. The intracellular
the presence of systemic inflammation was also studied in ex-
changes observed by exposing leukocytes of healthy volunteers
perimental models of sepsis and sepsis-induced ARDS (33, 37,
to plasma samples collected before randomization included
38). In a sheep model of sepsis-induced ARDS, maximal bind-
escalating increases in NF-B–mediated activities (NF-B
ing capacity of GR decreased continuously after endotoxin
DNA-binding, p50 and p65 DNA-binding, and transcription
infusion, whereas there was a marked elevation of cortisol
of TNF-␣ and IL-1) and modest changes in GR-␣–mediated
concentrations (37). The reduced GR binding correlated neg-
activities. The reduction in cytoplasmic IB␣ levels observed
atively (r ϭ Ϫ0.87; p Ͻ 0.01) with phospholipase A2 activity, a
before randomization indicates that NF-B–mediated IB␣
gene that is stimulated by NF-B. In a rat model of septic
degradation predominated over GR-␣–mediated IB␣ forma-
shock, GR blockade by mifepristone (RU 486) exacerbated
the physiologic and pathologic changes induced by endotox-
If acquired glucocorticoid receptor resistance played a role in
emia (38). Phospholipase A2 activity in rats with 80% GR
the pathogenesis of unresolving ARDS, adequate hormonal sup-
blockade was more marked than in those with 50% GR block-
plementation should restore glucocorticoid antiinflammatory
ade (38). Monocytes of patients with sepsis developed near-
function, by decreasing the production of inflammatory cyto-
total glucocorticoid resistance in vitro when cytokines, espe-
kines, cytokine-driven HPA-axis activity, and cytokine-driven
organ dysfunction. Indeed, after randomization, the biologic
Several inflammatory cytokines, including TNF-␣, IL-1,
and physiologic characteristics of the two groups (methylpred-
and IL-6, activate NF-B (39). It has been proposed that when
nisolone versus placebo) rapidly diverged. The responses ob-
cytokine-activated NF-B forms protein–protein complexes
served during methylprednisolone administration support the
with activated GR-␣, the availability and activity of effective
concept of inflammation-dependent acquired glucocorticoid
GR-␣ molecules are reduced (6, 31). This functional reduction
resistance in patients with ARDS. We found that the methyl-
in GR-␣ availability is associated with decreased GR-␣–GRE
prednisolone-treated group had significant, progressive reduc-
DNA binding and GC-mediated antiinflammatory activity (6,
tions in plasma concentrations of TNF-␣, IL-1, IL-6, ACTH,
AMERICAN JOURNAL OF RESPIRATORY AND CRITICAL CARE MEDICINE
Figure 7. Relations on nat- ural logarithmic scales be- tween nuclear NF-B and nuclear GR-␣ (top) and be- tween nuclear NF-B and cytoplasmic GR-␣ bound to NF-B (bottom). Mean in- tracellular changes observed by exposing PBL of a healthy volunteer to plasma sam- ples collected at random- ization (rand) and after 3, 5, 7, and 10 days in the methylprednisolone (open triangles) and placebo (open circles) groups. See text for explanation.
and cortisol and a parallel improvement in severity of organdysfunction scores (18). The biologic response observed in thisstudy was similar to that of a prior uncontrolled report (41). Inthat study, methylprednisolone treatment was associated withsignificant and parallel reductions in plasma and bronchoalve-olar lavage (BAL) TNF-␣, IL-1, IL-6, and IL-8 concentra-tions; LIS; and BAL indices of pulmonary vascular permeabil-ity (BAL albumin and percentage of polymorphonuclearcells) (41).
Our ex vivo findings reflect the effects of a mixture of in-
flammatory cytokines and other factors and the variable me-thylprednisolone and cortisol concentrations in the plasma ofour patients on normal nonactivated circulating blood leuko-
Figure 8. Relations on natural logarithmic scales between IB␣ and
cytes. The concentration of methylprednisolone in the plasma
factors affecting its formation (nuclear GR-␣) and degradation (IL-10
samples was unknown and may have varied during the study
mRNA and TNF-␣ mRNA). Mean intracellular changes observed by ex-
period. A prior study has shown similar changes in NF-B ac-
posing PBL of a healthy volunteer to plasma samples collected at ran-domization (rand) and after 3, 5, 7, and 10 days in the methylpred-
tivation in the peripheral monocytes of patients with septic
nisolone (open triangles) and placebo (open circles) groups. See text for
shock (42) or with critical illness (43). These data suggest that
the glucocorticoid resistance of the immune system of ARDSpatients simply reflects the inflammatory state of the individual.
The extent to which methylprednisolone in the plasma has
biased the magnitude of responses is unknown. We conducted
on the exposure of normal leukocytes to plasma samples from
an experiment to quantify this bias (data not reported). The
patients receiving methylprednisolone or placebo. With meth-
amount of IL-8 released from fresh PBL incubated for four
ylprednisolone administration, the intracellular relations be-
hours with plasma from a patient who received methylpred-
tween the NF-B and GR-␣ signaling pathways changed from
nisolone was four times greater when the methylprednisolone
an initial NF-B–driven and GR-␣–resistant state to a GR-␣–
had been removed by dialysis compared with undialyzed
driven and GR-␣–sensitive one. In contrast, exposure to serial
plasma. For TNF-␣ release, the effect of removing methyl-
plasma samples collected during placebo administration dem-
prednisolone was an increase of 1.6 times the amount compared
onstrated a significant—but lower (in comparison with the
with undialyzed plasma. Thus, the degree of bias probably
methylprednisolone group, p Ͻ 0.0001)—increase in GR-␣–
varies greatly, depending on what intracellular or extracellular
mediated activity over time, and persistently elevated NF-B
variable is selected. We point out that in the current report the
results from PBL of a healthy volunteer are consistent withthe outcomes of patients randomized in the clinical trial (18). Acknowledgment : The authors recognize James Dalton, Pharm.D., and
However, we explicitly acknowledge this known source of bias
Pierluigi Carratu, M.D., for the critical review of this manuscript. They are
on the magnitude, significance, and possibly the direction of
indebted to their research nurses, Stephanie Carson, Mary Gibson, andReba Umberger; to David Patterson for laboratory assistance; and to their
the differences presented in this report.
collaborating clinical investigators, Drs. Emmel Golden and Stacey A. Head-
Our current understanding of the physiologic antagonism
ley. They acknowledge Vivian Gomez for the creation of the figures, and
between the NF-B and GR-␣ cascades explains our findings
Gail Spake and David Armbruster for editing the manuscript.
Meduri, Tolley, Chrousos, et al.: Methylprednisolone Suppresses Inflammation in Unresolving ARDS
References
22. Slutsky AS. Mechanical ventilation: American College of Chest Physi-
cians’ Consensus Conference. Chest 1993;104:1833–1859.
1. Meduri GU, Headley S, Kohler G, Stentz F, Tolley E, Umberger R,
23. Bernard GR, Doig G, Hudson LD, Lemeshow S, Marshall JC, Russell J,
Leeper K. Persistent elevation of inflammatory cytokines predicts a poor
Sibbald W, Sprung CL, Vincent JL, Wheeler AP. Quantification of or-
outcome in ARDS: plasma IL-1 beta and IL-6 levels are consistent and
gan failure for clinical trials [abstract]. Am J Respir Crit Care Med
efficient predictors of outcome over time. Chest 1995;107:1062–1073.
2. Krafft P, Fridrich P, Pernerstorfer T, Fitzgerald RD, Koc D, Schneider B,
24. Meduri GU, Tolley EA, Chinn A, Stentz F, Postlethwaite A. Procol-
Hammerle AF, Steltzer H. The acute respiratory distress syndrome:
lagen types I and III aminoterminal propeptide levels during acute
definitions, severity and clinical outcome: an analysis of 101 clinical in-
respiratory distress syndrome and in response to methylpredniso-
vestigations. Intensive Care Med 1996;22:519–529.
lone treatment. Am J Respir Crit Care Med 1998;158(5 Pt 1):1432–
3. McKay LI, Cidlowski JA. Molecular control of immune/inflammatory
responses: interactions between nuclear factor-kappa B and steroidreceptor-signaling pathways. Endocr Rev 1999;20:435–459.
25. Kirk RE. Split-plot factorial design: design with group-treatment con-
4. Baeuerle PA, Baltimore D. NF-kappa B: ten years after. Cell 1996;
founding. Experimental design, 2nd ed. Wadsworth, Inc.: Belmont,
5. Barnes PJ, Karin M. Nuclear factor-kappa B: a pivotal transcription factor
26. Chrousos GP, Detera-Wadleigh SD, Karl M. Syndromes of glucocorti-
in chronic inflammatory diseases. N Engl J Med 1997;336:1066–1071.
coid resistance. Ann Intern Med 1993;119:1113–1124.
6. Bamberger CM, Schulte HM, Chrousos GP. Molecular determinants of
27. Lamberts SW, Koper JW, Biemond P, den Holder FH, de Jong FH. Cor-
glucocorticoid receptor function and tissue sensitivity to glucocorti-
tisol receptor resistance: the variability of its clinical presentation and
coids. Endocr Rev 1996;17:245–261.
response to treatment. J Clin Endocrinol Metab 1992;74:313–321.
7. Didonato JA, Saatcioglu F, Karin M. Molecular mechanisms of immuno-
28. Lane SJ, Lee TH. Glucocorticoid receptor characteristics in monocytes
suppression and anti-inflammatory activities by glucocorticoids. Am J
of patients with corticosteroid-resistant bronchial asthma. Am RevRespir Crit Care Med 1996;154:S11–S15. Respir Dis 1991;143(5 Pt 1):1020–1024.
8. Scheinman RI, Cogswell PC, Lofquist AK, Baldwin AS Jr. Role of tran-
29. Sher ER, Leung DY, Surs W, Kam JC, Zieg G, Kamada AK, Szefler SJ.
scriptional activation of I kappa B alpha in mediation of immunosup-
Steroid-resistant asthma: cellular mechanisms contributing to inadequate
pression by glucocorticoids. Science 1995;270:283–286.
response to glucocorticoid therapy. J Clin Invest 1994;93:33–39.
9. Wissink S, van Heerde EC, vand der Burg B, van der Saag PT. A dual
30. Adcock IM, Lane SJ, Brown CR, Peters MJ, Lee TH, Barnes PJ. Differ-
mechanism mediates repression of NF-kappa B activity by glucocorti-
ences in binding of glucocorticoid receptor to DNA in steroid-resis-
coids. Mol Endocrinol 1998;12:355–363.
tant asthma. J Immunol 1995;154:3500–3505.
10. Wang P, Wu P, Siegel MI, Egan RW, Billah MM. Interleukin (IL)-10 in-
31. Barnes PJ, Greening AP, Crompton GK. Glucocorticoid resistance in
hibits nuclear factor kappa B (NF kappa B) activation in human
asthma. Am J Respir Crit Care Med 1995;152(6 Pt 2):S125–S140.
monocytes: IL-10 and IL-4 suppress cytokine synthesis by different
32. Norbiato G, Bevilacqua M, Vago T, Baldi G, Chebat E, Bertora P,
mechanisms. J Biol Chem 1995;270:9558–9563.
Moroni M, Galli M, Oldenburg N. Cortisol resistance in acquired
11. Hofmann TG, Hehner SP, Bacher S, Droge W, Schmitz ML. Various
immunodeficiency syndrome. J Clin Endocrinol Metab 1992;74:
glucocorticoids differ in their ability to induce gene expression, apop-
tosis and to repress NF-kappaB-dependent transcription. FEBS Lett
33. Molijn GJ, Spek JJ, van Uffelen JC, de Jong FH, Brinkmann AO, Bruin-
ing HA, Lamberts SW, Koper JW. Differential adaptation of gluco-
12. Sheppard KA, Phelps KM, Williams AJ, Thanos D, Glass CK, Rosen-
corticoid sensitivity of peripheral blood mononuclear leukocytes in
feld MG, Gerritsen ME, Collins T. Nuclear integration of glucocorticoid
patients with sepsis or septic shock. J Clin Endocrinol Metab 1995;80:
receptor and nuclear factor- kappaB signaling by CREB-binding protein
and steroid receptor coactivator-1. J Biol Chem 1998;273:29291–29294.
34. Almawi WY, Lipman ML, Stevens AC, Zanker B, Hadro ET, Strom TB.
13. Melby JC, Spink WW. Comparative studies on adrenalcortical function
Abrogation of glucocorticoid-mediated inhibition of T cell prolifera-
and cortisol metabolism in healthy adults and in patients with shock
tion by the synergistic action of IL-1, IL-6, and IFN-gamma. J Immu-
due to infection. J Clin Invest 1958;37:1791–1798.
14. Reincke M, Allolio B, Wurth G, Winkelmann W. The hypothalamic-
35. Kam JC, Szefler SJ, Surs W, Sher ER, Leung DY. Combination IL-2 and
pituitary-adrenal axis in critical illness: response to dexamethasone and cor-
IL-4 reduces glucocorticoid receptor-binding affinity and T cell re-
ticotropin-releasing hormone. J Clin Endocrinol Metab 1993;77:151–156.
sponse to glucocorticoids. J Immunol 1993;151:3460–3466.
15. Briegel J, Forst H, Hellinger H, Haller M. Contribution of cortisol defi-
36. Spahn JD, Szefler SJ, Surs W, Doherty DE, Nimmagadda SR, Leung DY.
ciency to septic shock. Lancet 1991;338:507–508.
A novel action of IL-13: induction of diminished monocyte glucocorti-
16. Annane D, Sebille V, Troche G, Raphael JC, Gajdos P, Bellissant E. A
coid receptor-binding affinity. J Immunol 1996;157:2654–2659.
3-level prognostic classification in septic shock based on cortisol levels
37. Liu LY, Sun B, Tian Y, Lu BZ, Wang J. Changes of pulmonary gluco-
and cortisol response to corticotropin. JAMA 2000;283:1038–1045.
corticoid receptor and phospholipase A2 in sheep with acute lung in-
17. Meduri GU, Chrousos GP. Duration of glucocorticoid treatment and
jury after high dose endotoxin infusion. Am Rev Respir Dis 1993;148:
outcome in sepsis: is the right drug used the wrong way? Chest 1998;
38. Fan J, Gong XQ, Wu J, Zhang YF, Xu RB. Effect of glucocorticoid re-
18. Meduri GU, Headley S, Carson S, Umberger R, Kelso T, Tolley E. Pro-
ceptor (GR) blockade on endotoxemia in rats. Circ Shock 1994;42:76–82.
longed methylprednisolone treatment improves lung function and
39. Baiuerle PA, Baichwal VR. NF-kappa B as a frequent target for immu-
outcome of unresolving ARDS: a randomized, double-blind, placebo-
nosuppressive and anti-inflammatory molecules. Adv Immunol 1997;
controlled trial. JAMA 1998;280:159–165.
19. Meduri GU. An historical review of glucocorticoid treatment in sepsis:
40. Cato AC, Wade E. Molecular mechanisms of anti-inflammatory action
disease pathophysiology and the design of treatment investigation.
of glucocorticoids. Bioessays 1996;18:371–378.
41. Meduri GU, Headley S, Tolley E, Shelby M, Stentz F, Postlethwaite A.
20. Bernard GR, Artigas A, Brigham KL, Carlet J, Falke K, Hudson L,
Plasma and BAL cytokine response to corticosteroid rescue treatment
Lamy M, Legall JR, Morris A, Spragg R. The American-European
in late ARDS. Chest 1995;108:1315–1325.
Consensus Conference on ARDS: definitions, mechanisms, relevant
42. Bohrer H, Qiu F, Zimmermann T, Zhang Y, Jllmer T, Mannel D, Botti-
outcomes, and clinical trial coordination. Am J Respir Crit Care Med
ger BW, Stern DM, Waldherr R, Saeger HD, et al. Role of NF kappa
B in the mortality of sepsis. J Clin Invest 1997;100:972–985.
21. Murray JF, Matthay MA, Luce JM, Flick MR. An expanded definition
43. Paterson RL, Galley HF, Dhillon JK, Webster NR. Increased nuclear
of the adult respiratory distress syndrome. Am Rev Respir Dis 1988;
factor kB activation in critically ill patients who die. Crit Care Med
GROWTH OF EIGHT-YEAR-OLD AMERICAN GINSENG IN A RED MAPLE FOREST AS INFLUENCED BY LIME AND ORGANIC FERTILIZER APPLICATION Alain Olivier, Isabelle Nadeau, Hakim Ouzennou, Justin Panadu Dzaringa, Department of Plant Science, Université Laval, Quebec City (Qc), Canada ABSTRACT In Québec, the suitability of nutrient-depleted, acidic soils of red maple ( Acer rubrum L.) forests t
Schroder USA Fondo Común de Inversión Estados Contables correspondientes al período de nueve meses finalizado el 30 de septiembre de 2009 presentados en forma comparativa. Schroder USA Fondo Común de Inversión Estados Contables correspondientes al período de nueve meses finalizado el 30 de septiembre de 2009 presentados en forma comparativa. Indice Balance Genera
 Meduri, Tolley, Chrousos, et al.: Methylprednisolone Suppresses Inflammation in Unresolving ARDS
and up to randomization, plasma ACTH and cortisol levelswere similar in both groups. After randomization, the methyl-
Cytokine Concentrations from Plasma of Randomized Patients
Meduri, Tolley, Chrousos, et al.: Methylprednisolone Suppresses Inflammation in Unresolving ARDS
and up to randomization, plasma ACTH and cortisol levelswere similar in both groups. After randomization, the methyl-
Cytokine Concentrations from Plasma of Randomized Patients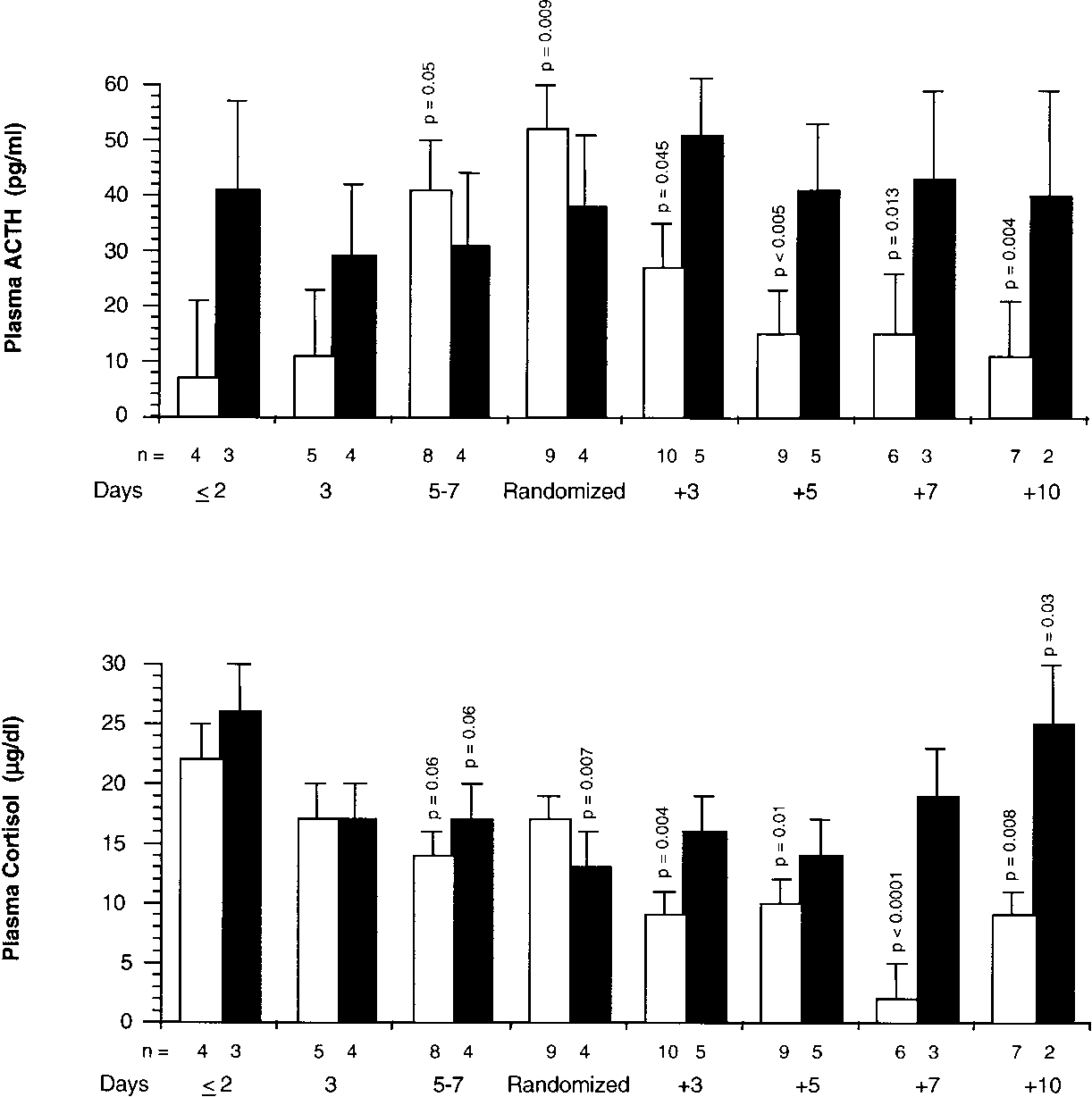 AMERICAN JOURNAL OF RESPIRATORY AND CRITICAL CARE MEDICINE
Figure 2. Plasma ACTH and cortisol levels before and after
AMERICAN JOURNAL OF RESPIRATORY AND CRITICAL CARE MEDICINE
Figure 2. Plasma ACTH and cortisol levels before and after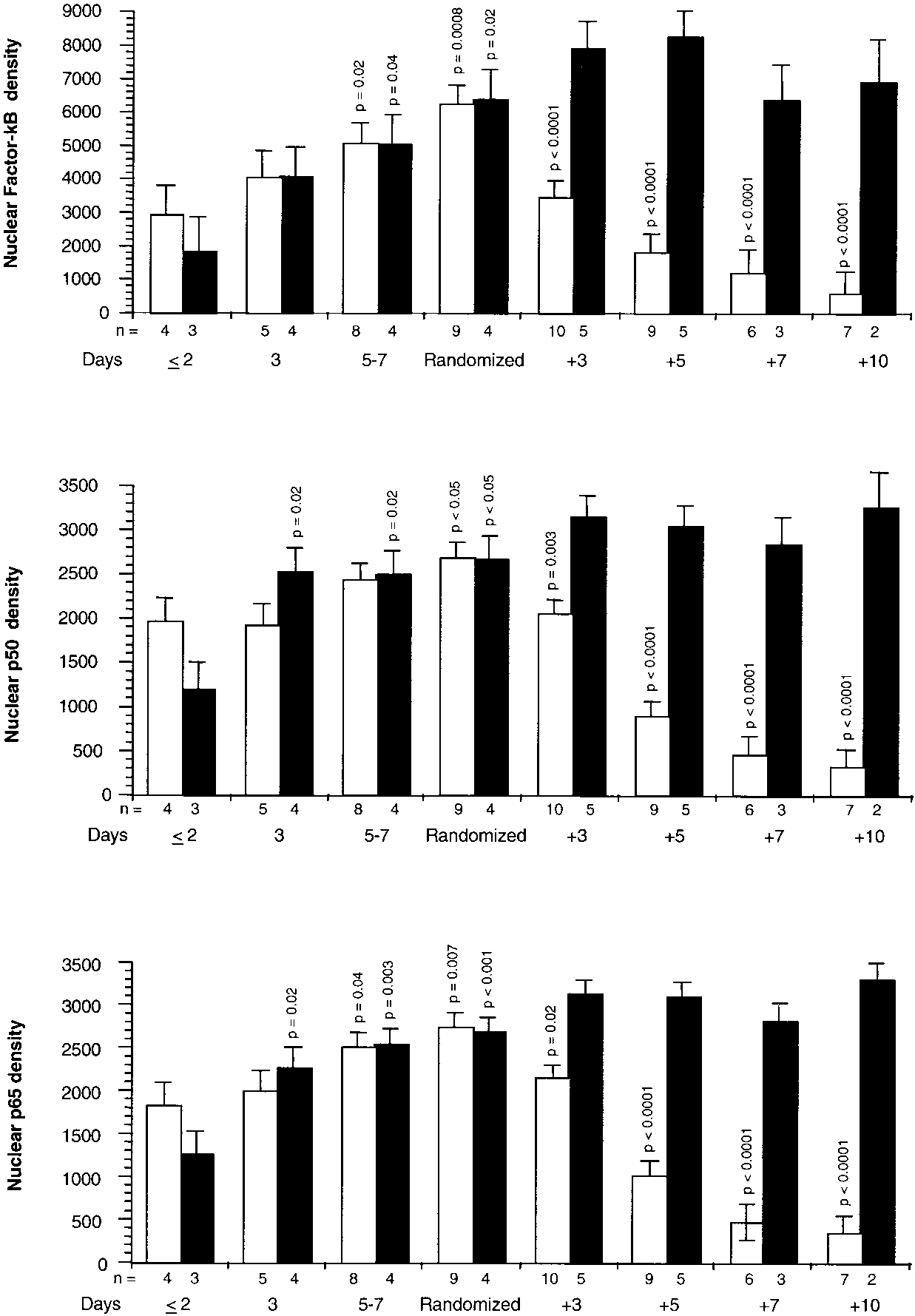 Meduri, Tolley, Chrousos, et al.: Methylprednisolone Suppresses Inflammation in Unresolving ARDS
Figure 3. NF-B, p65 subunit, and p50 subunit binding to
Meduri, Tolley, Chrousos, et al.: Methylprednisolone Suppresses Inflammation in Unresolving ARDS
Figure 3. NF-B, p65 subunit, and p50 subunit binding to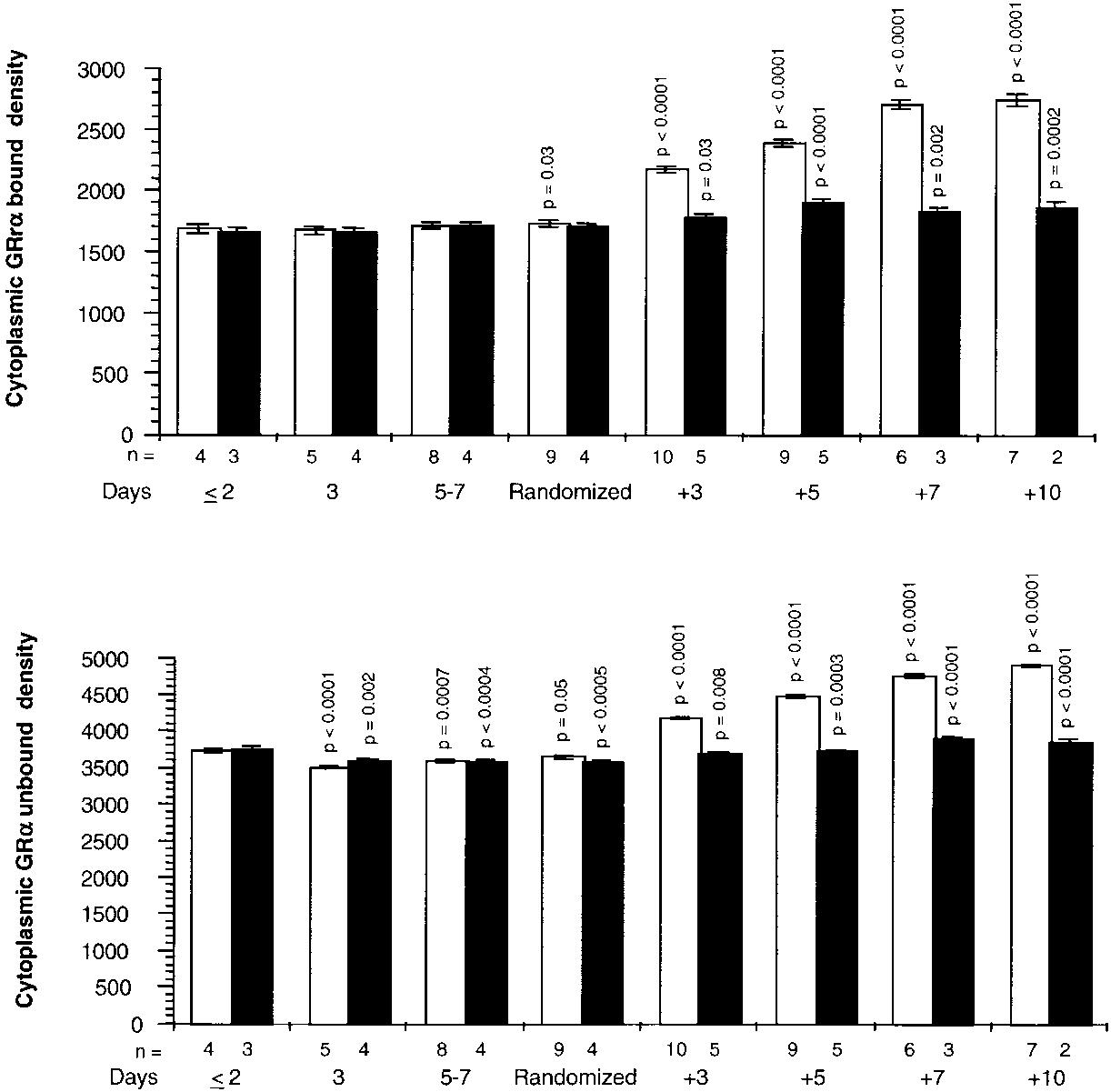
 AMERICAN JOURNAL OF RESPIRATORY AND CRITICAL CARE MEDICINE
Figure 4. Cytoplasmic GR bound and unbound to NF-B
AMERICAN JOURNAL OF RESPIRATORY AND CRITICAL CARE MEDICINE
Figure 4. Cytoplasmic GR bound and unbound to NF-B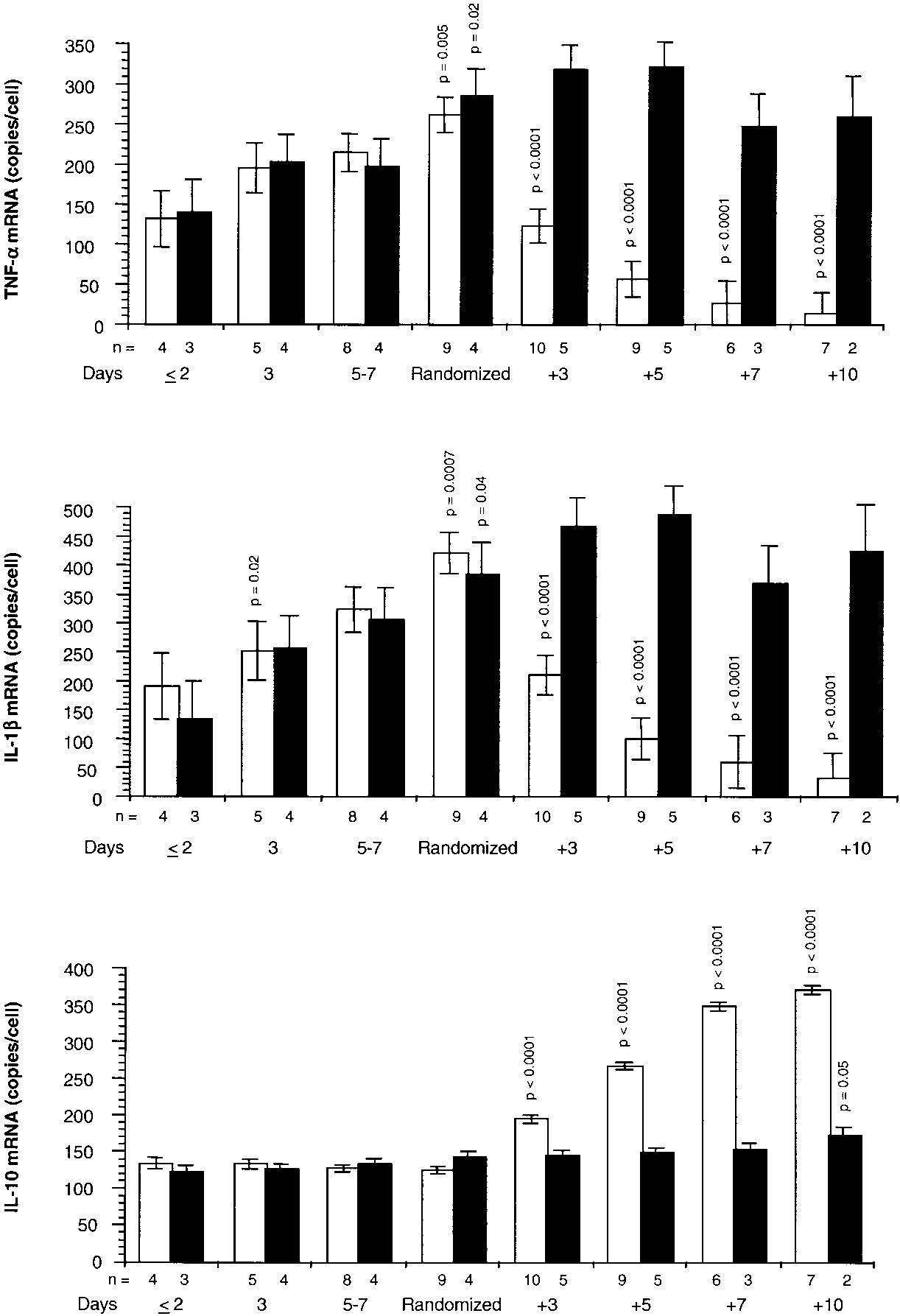 Meduri, Tolley, Chrousos, et al.: Methylprednisolone Suppresses Inflammation in Unresolving ARDS
Figure 6. Messenger RNA TNF-␣, IL-1, and IL-10 before
Meduri, Tolley, Chrousos, et al.: Methylprednisolone Suppresses Inflammation in Unresolving ARDS
Figure 6. Messenger RNA TNF-␣, IL-1, and IL-10 before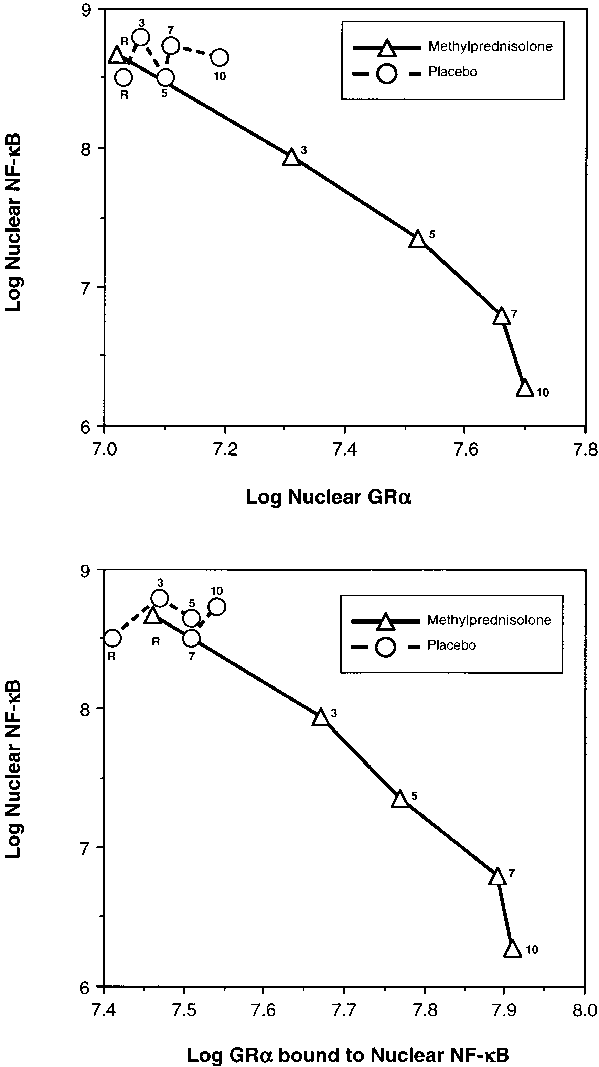
 AMERICAN JOURNAL OF RESPIRATORY AND CRITICAL CARE MEDICINE
Figure 7. Relations on nat-
AMERICAN JOURNAL OF RESPIRATORY AND CRITICAL CARE MEDICINE
Figure 7. Relations on nat-