Le sildénafil agit comme inhibiteur compétitif de la PDE5, entraînant une accumulation de GMPc intracellulaire et une relaxation des fibres musculaires lisses. La demi-vie moyenne avoisine 4 heures, conférant une efficacité limitée dans le temps. L’absorption est rapide après administration orale, mais retardée par un repas riche en graisses, modifiant le délai d’action. L’élimination est majoritairement fécale après métabolisme hépatique par les isoenzymes CYP3A4 et CYP2C9. Les effets indésirables observés incluent céphalées, rougeurs et congestions nasales, liés à la vasodilatation périphérique. Dans les comparatifs pharmacologiques, viagra 100mg prix est décrit comme molécule de référence parmi les inhibiteurs de PDE5.
Pii: s0928-4257(00)00154-6
J. Physiol. (Paris) 94 (2000) 135−138 2000 Elsevier Science Ltd. Published by Éditions scientifiques et médicales Elsevier SAS. All rights reservedS0928425700001546/FLA
Modulation by GABA and delta opioid receptors of neurally induced responses in isolated guinea-pig taenia coli and human colonic circular muscle
Éva Makóa,b*, András Z Rónaia, György Ádámb, Gábor Juhászb,
László Ritterc, Béla Lestárc, Vincenzo Crunellid
aDepartment of Pharmacology, Semmelweis University of Medicine, P.O.B. 370, 1445 Budapest, HungarybDepartment of Comparative Physiology, Eötvös University, Budapest, HungarycDepartment of Surgery, Haynal Postgraduate University of Medicine, Budapest, HungarydUniversity of Wales, College of Cardiff, Cardiff, UKAbstract — The GABA-ergic and opioid modulation of neurally induced muscle responses was studied in isolated guinea-pig taenia coli and human colonic circular muscle, using identical field stimulation parameters (rectangular pulses of 0.5 ms duration, 9 V⋅cm–1 intensity, trains of 3 pulses at 0.5 Hz, repeated every 1/3/5 min).The stimulation-induced contractions were inhibited in both preparations by GABA and baclofen; the IC
values in human colonic circular muscle were z100 and 31.0 µM, respectively. In
guinea-pig taenia coli, the inhibition by 10–4 M GABA was dose-dependently reversed by 10–4–10–3 M of GABA receptor antagonist
CGP 35348; antagonism by phaclofen was less effective in the same concentration range. In human colonic circular muscle, inhibitionby 3 × 10–5 M baclofen was fully reversed by 10–3 M CGP 35348. With the exception of caecum, the delta 2 opioid receptor agonistdeltorphin II was a potent inhibitor in human colonic circular muscle. 10–8 M Deltorphin caused a 74.4 ± 9.6% (n = 4) inhibition whichwas reversed by 10–6 M of delta receptor selective peptide antagonist BOC-Tyr-Pro-Gly-Phe-Leu-Thr(OtBu). Deltorphin II wasineffective in guinea-pig taenia coli even at 10–6 M; the same concentration caused an 84.3 ± 7.9 (n = 4) inhibition in humanpreparations. It is concluded that: 1) GABA-ergic modulatory mechanisms are present both in human colonic circular muscle andguinea-pig taenia coli; 2) the GABA receptors involved are of type B; and 3) delta opioid receptor-mediated modulation functions onlyin human colonic circular muscle in regions other than the caecum. 2000 Elsevier Science Ltd. Published by Éditions scientifiqueset médicales Elsevier SAS
guinea-pig taenia coli / human colonic circular muscle / GABA receptors / delta opioid receptors 1. Introduction
lonic circular muscle ([9] and Makó in preparation). Inthe human colonic circular muscle, 5HT /H receptor
mediated link is likely to be present in the stimulatory
proenkephalinA/prodynorphin-derived opioid peptides
neural network since cyproheptadine inhibited the
are neurotransmitters of enteric interneurons and com-
neural stimulation-induced contractions ([9] and Makó
prise part of the intrinsic neural circuits regulating
in preparation). To compare the opioid and GABA-
peristalsis [1, 2, 5, 6, 8]. Our study was aimed at
ergic modulatory mechanisms in the two isolated
characterising the receptor types where these transmit-
organs, the delta (delta 2) opioid receptor agonist
ters exert their action in isolated, neurally stimulated
deltorphin II (DT-II), the delta receptor selective pep-
human colonic circular muscle and guinea-pig taenia
tide antagonist BOC-Tyr-Pro-Gly-Phe-Leu-Thr(OtBu)
coli. The common technical motif for both prepara-
[BOC-YPGFLT(OtBu)] [12], GABA, the GABA re-
tions was the choice of parameters of field neural
stimulation. These parameters (short trains at a low
antagonist CGP 35348 and phaclofen [3, 4, 7, 10] were
frequency) were expected to facilitate the detection
both of inhibitory and stimulatory actions of exog-enously administered receptor-specific agents. Thechosen field stimulation parameters elicited a mul-
2. Materials and methods
tiphasic or apparently monophasic muscle response
Drugs used were as follows: (-) baclofen (Research
with a dominant contractile component in both isolated
Biochemicals International), CGP 35348 (Tocris
organs ([9, 11] and Makó in preparation). The neural
Neuramin), gamma-amino butyric acid (Sigma),
circuitry involved in the contractile responses of the
phaclofen (Tocris Neuramin). BOC-Tyr-Pro-Gly-Phe-
two preparations is, naturally, quite different. The
Leu-Thr(OtBu) was synthesised at the Research Group
contractions were atropine-sensitive in guinea-pig tae-
of Peptide Chemistry, Eötvös University, Budapest, as
nia coli [11] whereas atropine-resistant in human co-
described previously [12]. Deltorphin II (Tyr-D-Ala-Phe-Glu-Val-Val-Gly-NH ) was kindly supplied by G.
Tóth of Isotope Laboratory, Biological Research Cen-
tre of Hungarian Academy of Sciences, Szeged. All theother substances were of analytical grade.
Male guinea-pigs, weighing 250–350 g, were used
and the taenia coli segments were prepared andmounted as described previously [11]. In brief,5–8 mm long segments were mounted in Krebs’ solu-tion aerated with carbogen (O :CO = 95:5) at 37 °C.
An initial tension of 0.5 g was applied re-adjusted onceafter 5 min of 45–60 min equilibration period.
Subjects were 8 patients, 4 male and 4 female, age
28–76 years, undergoing colonic surgery for cancer (6)colitis ulcerosa (1) or slow colon (1). The preparationswere made from non-diseased tissue dissected accord-ing to the pre-set rules of disease-dependent surgical
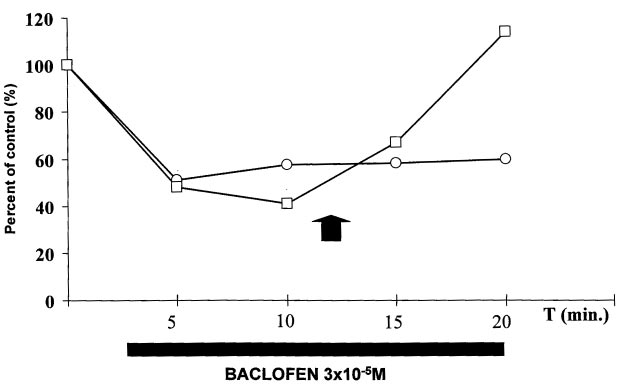
Figure 1. The inhibition of neural field stimulation-induced
protocols. Among the dissected tissues, the sigmoid
contractions of human colonic circular muscle by baclofen
colon (4), the rectum (4) and the caecum (1) were
and the reversal of inhibition by CGP 35348. ·, 3 × 10–5 M
represented (i.e. from one subject, two specimens were
baclofen alone; [, 3 × 10–5 M baclofen + 10–3 M CGP35348
dissected). The postoperative use of tissue samples
added later (arrow). Points represent the mean of two
was approved by the Ethical Committee of the Uni-
versity. The routinely used preoperative medicationwas metronidazole and aminoglycoside as antimicro-bials and benzodiazepine as sedative. Five operations
3. Results
were performed under general anesthesia; in threecases, epidural anesthesia was used.
Of the total of 65 human colonic circular muscle
The specimens were transported in a special cham-
strips, 61 responded to neural field stimulation. Only
ber in thermostated (33 °C) carbogenated Krebs’ solu-
those preparations were used for the pharmacological
tion; the transportation took less than 20 min. The
tests where an apparently monophasic contractile re-
preparation was done at room temperature in carboge-
sponse was present with or without spontaneous back-
nated Krebs’ solution. After the removal of the mu-
ground activity. Both GABA and baclofen inhibited
cosal and serosal layer, 10–15 mm long, 2–2.5 mm
the evoked contractions; in the case of the former,
wide circularly oriented strips were cut from the
there was a high propensity to develop tachyphylaxis.
intertaenial segment. The strips were mounted in
At 10–4 M concentration, GABA exerted 55.4 ± 18.3%
carbogenated Krebs’ solution at 37 °C. The muscles
(mean ± SEM, n = 3) inhibition; at 10–3 M concentra-
were left without applying any tension for 15 min,
tion, the inhibition tended to be less than 50%.
washed once, then 1-g tension was applied. The
Baclofen had a dose-dependent inhibitory effect with
tension was re-adjusted after 10 min then the prepara-
only a minor tendency for tachypylaxis; the IC
tions were equilibrated for 60 min under resting con-
was 31.0 µM (28.6, 33.4, n = 2). The inhibition caused
ditions. The composition of Krebs’ solution was as
by 3 × 10–5 M baclofen could be reversed by the
follows (in mmol⋅L–1): NaCl 118.0, NaHCO 25.0,
GABA receptor antagonist CGP 35348 (figure 1).
KCl 4.7, KH PO 1.2, CaCl 2.5, MgSO 1.2 (human)
According to our previous findings in guinea-pig
= 4.5 ± 1.0 µM, n = 5) but not mus-
The parameters of field stimulation [11] were: rect-
cimol (at 10–4 M) had inhibitory effect in preparations
angular impulses of 0.5 ms duration, 9 V⋅c–1m (supra-
stimulated neurally by parameters closely matching
maximal) intensity; trains of 3 pulses at 0.5 Hz fre-
the ones used presently. We characterised further this
quency were repeated by 1 (guinea-pig) or 5 min
inhibitory action by the reversal of the inhibition by
(human). The contractions were measured under non-
10–4 M GABA by cumulatively administered GABA
isometric conditions. From a single surgical specimen,
receptor antagonists CGP 35348 and phaclofen (fig-
4–12 strips were prepared; they were randomly as-
signed to the different pharmacological interventions.
Besides GABA and baclofen, the delta 2 opioid
In one preparation, only a single drug or drug combi-
receptor agonist deltorphin II was also a potent inhibi-
tor in neurally stimulated human colonic circular
For statistical comparisons ANOVA followed by
muscle strips (figure 3) prepared from the sigmoid
Student’s t-test or just Student’s t-test was used,
colon (3 patients) or rectum (2 patients) but not in
depending on the experimental paradigms.
preparations made from the caecum (1 patient). Figure 4. The reversal of the inhibitory action of deltorphin Figure 2. The inhibition of neural field stimulation-induced
II by BOC-Tyr-Pro-Gly-Phe-Leu-Thr(OtBu) in neurally
contractions of guinea-pig taenia coli by GABA and the
stimulated human colonic circular muscle. Bars and vertical
reversal of inhibition by CGP 35348 or phaclofen. ·,
lines represent the mean ± SEM of 4 independent experi-
10–4 M GABA alone; [, 10–4 M GABA + 10–4, 3 × 10–4 and
10–3 M CGP 35348 added in a cumulative manner (arrows);
_, 10–4 M GABA + 10–4, 3 × 10–4 and 10–3 M phaclofen. Points and vertical lines represent the mean ± SEM of
by 10–8 M DT-II could be readily reversed by the delta
4 independent experiments. The reversal by CGP 35348 was
opioid receptor selective peptide antagonist BOC-
significant at 10–4 M and onward (ANOVA and Student’s
t-test) whereas for phaclofen it became significant only at10–3 M.
In circular muscle strips prepared from the caecum,
DT-II did not affect neural stimulation-induced con-tractions even at 10–6 M (n = 3, not shown); in sigmoid
The washout was very slow and in most cases
colon-derived strips obtained from the same patient,
incomplete therefore in one preparation only a single
DT-II was fully effective. Likewise, 10–6 M DT-II was
concentration of DT-II could be tested. There was an
ineffective in guinea-pig taenia coli (n = 4, not shown).
84.3 ± 7.9% (n = 4) inhibition at 10–6 M, 96.3 ± 3.0%(n = 3) at 10–7 M and 74.4 ± 9.6% (n = 4) at 10–8 M
4. Discussion
indicating that at these concentrations the effects fellinto the ‘plateau’ phase of the dose-response curve and
Two types of inhibitory modulation was studied in
must be below 10–8 M. The inhibition caused
two isolated organ preparations where the field-stimulation induced contractions have been shown tobe either atropine-, (guinea-pig taenia coli, [11]) ortetrodotoxin
muscle, [9] and Makó in preparation) indicating theinvolvement of neural elements. It was shown pres-ently that the GABA-ergic inhibitory modulation de-tected both in neurally stimulated human coloniccircular muscle and guinea-pig taenia coli is mediatedby B type of GABA receptors since 1.The action ofGABA is mimicked by the GABA receptor agonist
baclofen; 2.the effect of either GABA or baclofencould be reversed by the GABA receptor antagonist
CGP 35348. Of the GABA receptor antagonists we
phaclofen. In the CNS the localisation of CGP 35348sensitive GABA receptors has been characterised as
preferentially „hetero-presynaptic» relative to GABA-
Figure 3. The inhibitory effect of deltorphin II on two types
ergic neurons [3, 10] i.e. they are present on the
of neurally induced responses in human colonic circular
terminals of non-GABA-ergic neurons. The presence
muscle. Upper trace: inhibition of monophasic contractions
of GABA in mammalian enteric neural elements is
by deltorphin II (DT-II) and the reversal of inhibitionby BOC-Tyr-Pro-Gly-Phe-Leu-Thr(OtBu) (BOC-YPGFLT
extensively documented (e.g. [6]); in a subpopulation
(OtBu). Lower trace: inhibition of polyphasic contractions.
of human colonic neurons it has recently been de-
scribed to colocalise with enkephalin or nitric oxide
Froestl W., Mickel S.J., Von Sprecher G., Bittiger H., Olpe
synthase [8]. If the enteric localisation of CGP 35348
sensitive receptors follows the CNS pattern (i.e.theyare hetero-presynaptic) we may assume either facilita-
Hoyle C.H.V., Kamm M.A., Burnstock G., Lennard-Jones
tory GABA receptors at nerve terminals releasing an
J.E., Enkephalins modulate inhibitory neuromuscular trans-
mission in circular muscle of human colon via δ-opioid
inhibitory mediator, or inhibitory GABA receptors at
receptors, J. Physiol. 431 (1990) 465–478.
nerve terminals releasing an excitatory mediator.
Deltorphin II was a potent inhibitor of neurally
Jessen K.R., Hills J.M., Saffrey M.J., Immunohistochemical
mediated contractions in circular muscle strips from
demonstration of GABA-ergic neurons in the enteric nervoussystem, J. Neurosci. 6 (1986) 1628–1634.
human sigmoid colon or rectum but not from caecum;likewise, it was ineffective in guinea-pig taenia coli.
Kerr D.I., Ong J., Prager R.H., Gynther B.D., Curtis D.R.,
Delta and kappa opioid receptors are known to be
Phaclofen: a peripheral and central baclofen antagonist, BrainRes. 405 (1987) 150–154.
present in human colon [1, 5]. Altough deltorphin II isa selective agonist of delta 2 subtype of opioid
Krantis A., Nichols K., Staines W., Neurochemical character-
receptors in CNS tests [13], from the present results
ization and distribution of enteric GABA-ergic neurons andnerve fibres in the human colon, J. Auton. Nerv. Syst. 68
we cannot conclude that delta 2 receptors are present
in human colonic circular muscle. First, hitherto wehave been unable to detect delta 1/delta 2 opioid
Makó É., Rónai A.Z., Ádám Gy., Ritter L., Lestár B., In vitropharmacological analysis of the responses of human colonic
circular muscle to neural stimulation, in: 6th Joint Meeting of
nerve/smooth muscle preparations (Rónai, unpub-
the Italian, Hungarian and Polish Pharmacological Societies,
lished) nor are we aware of such successful differen-
tiation from other sources. Second, detailed further
[10] Raiteri M., Bonanno G., Paudice P., Cavazzani P., Schmid G.,
pharmacodynamic analysis is needed to make such a
Human brain cholecystokinin: release of cholecystokinin-like
declaration for the human colonic circular muscle; at
immunoreactivity (CCK-LI) from isolated cortical nerve end-
present the only valid statement is that these delta
ings and its modulation through GABA(B) receptors, J.
receptors are highly responsive to the agonist action of
Pharmacol. Exp. Ther. 278 (1996) 747–751.
[11] Rónai A.Z., Kardos J., Simonyi M., Potent inhibitory GABAB
receptors in stimulated guinea-pig taenia coli, Neuropharma-cology 26 (1987) 1623–1628. References
[12] Rónai A.Z., Magyar A., Orosz G., Borsodi A., Benyhe S., Tóth
G., Makó É., Kátay E., Babka E., Medzihradszky K., Opioid
Chamouard P., Klein A., Martin E., Adloff M., Angel F.,
antagonist properties of the highly δ-receptor selective BOC-
Regulatory role of enteric kappa opioid receptors in human
Tyr-Pro-Gly-Phe-Leu-Thr (OtBu) peptide and of its Phe1 and
colonic motility, Life Sci. 53 (1993) 1149–1156.
Mel1 analogues, Arch. Int. Pharmacodyn. 330 (1995)
Chamouard P., Rohr S., Meyer C., Baumann R., Angel F.,
Delta-opioid receptor agonists inhibit neuromuscular trans-
[13] Zaki A.P., Bilsky E.J., Vanderah T.W., Lai J., Evans C.J.,
mission in human colon, Eur. J. Pharmacol. 262 (1994) 33–39.
Porreca F., Opioid receptor types and subtypes: The δ receptor
Emri Z., Turner J.P., Crunelli V., Tonic activation of presyn-
as a model, Annu. Rev. Pharmacol. Toxicol. 36 (1996)
aptic GABA receptors on thalamic sensory afferents, Neu-
C. K. Gunsalus 217.333.1416 phone 244.1478 fax ! Current Positions: Director, National Center for Professional and Research Ethics Research Professor, Coordinated Science Laboratory ! Professional Experience Law, Ethics, Leadership and Professionalism ! Licensed to Practice Law in the State of Illinois Coordinated Science Laboratory, College of Engineering, University of Illinois!
SECURITY DETAILS Complying with de European Committee 91/155/EEC, last modification 2001/58/EC (07/27/01) and R.D. 255/2003. 1. IDENTIFICATION OF THE SUBSTANCE/PREPARATION AND OF THE COMPANY. VINFERMATON (SPECIAL COCKROACHES) P roduct Code: LABORATORIES VINFER S.A. POLIGONO IND’L CAMPOLLANO C/D Nº 2 TEL: 967 52 35 01 FAX. 967 24 29 14 +34 967 52 35 01
 tre of Hungarian Academy of Sciences, Szeged. All theother substances were of analytical grade.
tre of Hungarian Academy of Sciences, Szeged. All theother substances were of analytical grade.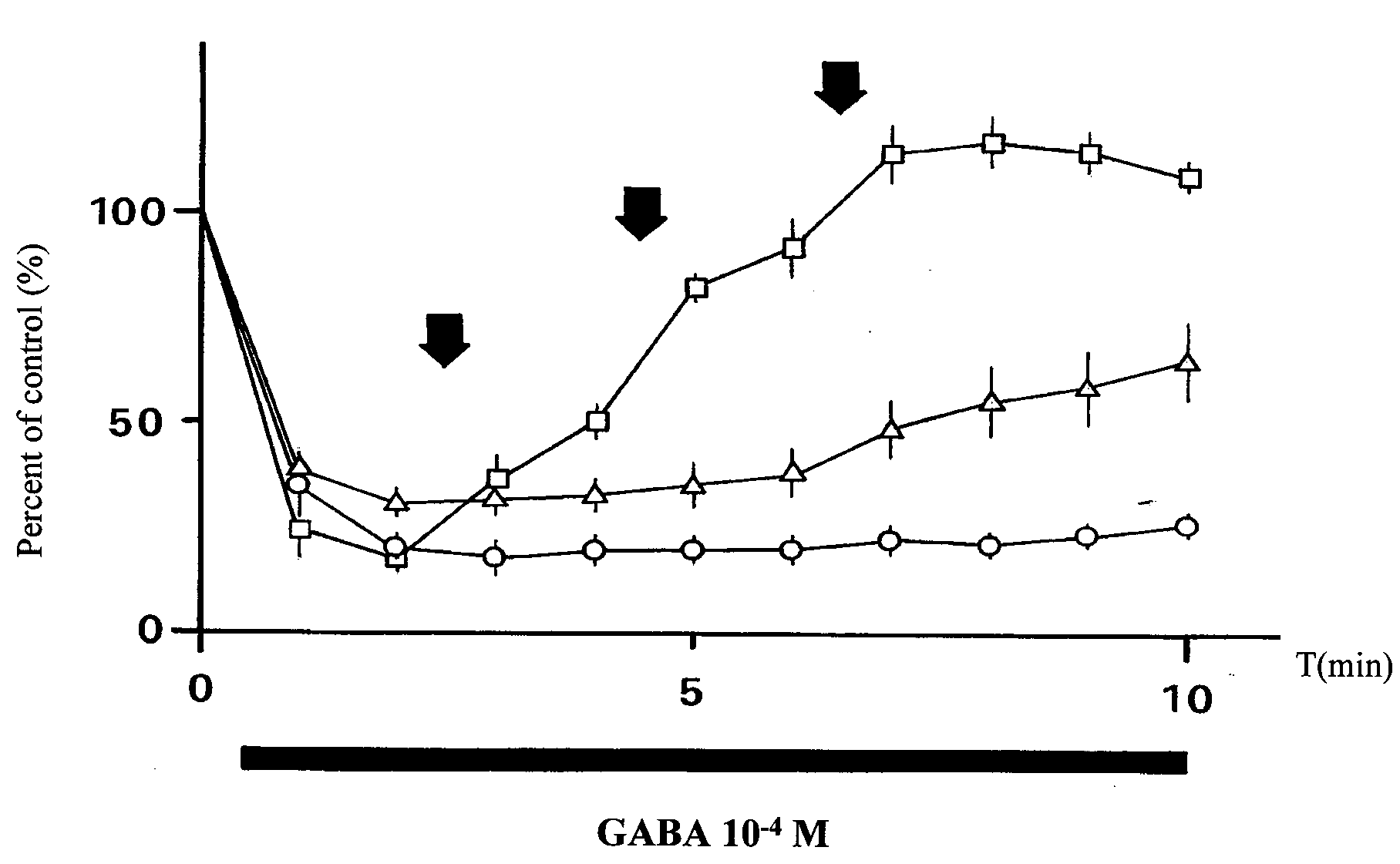
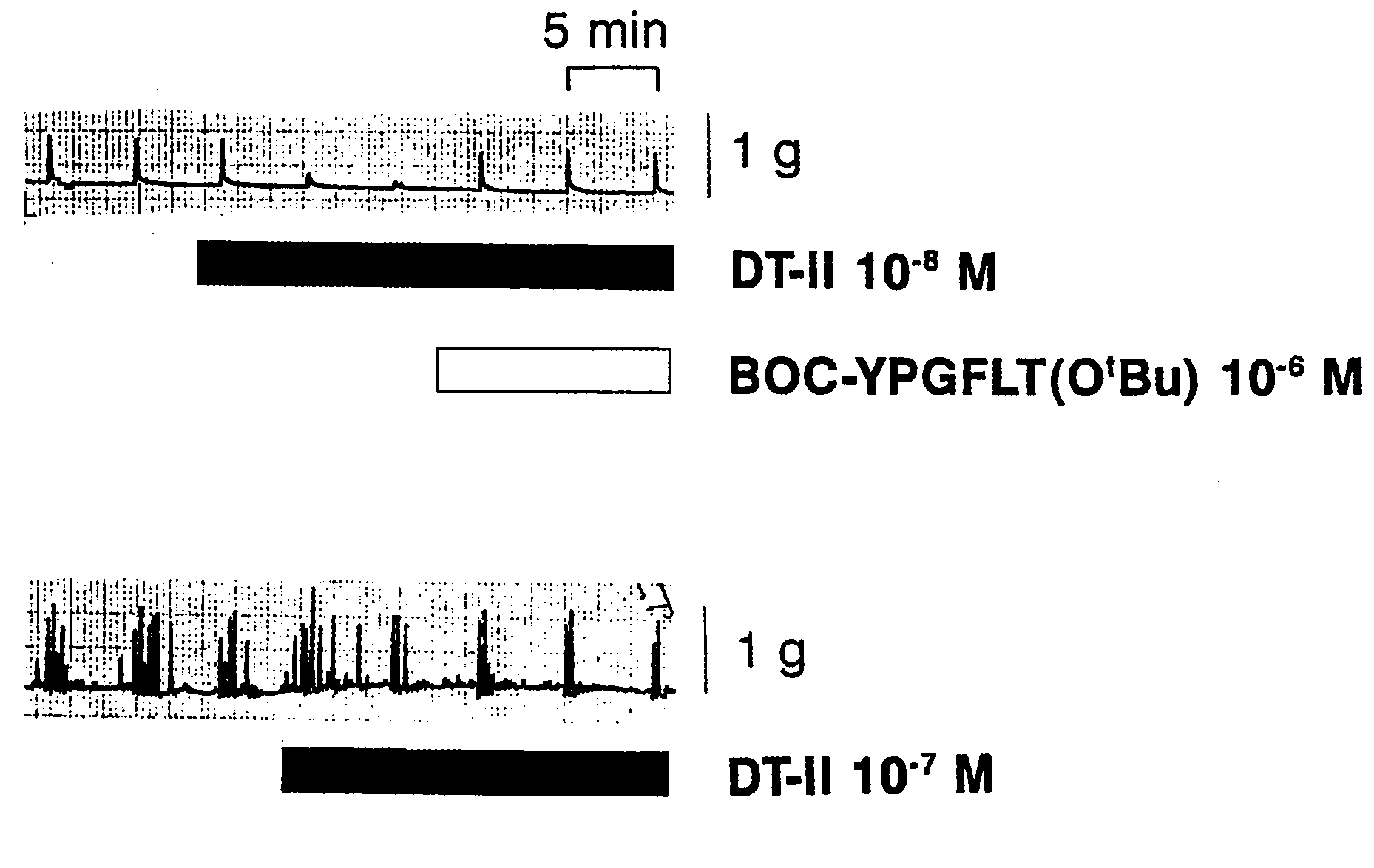
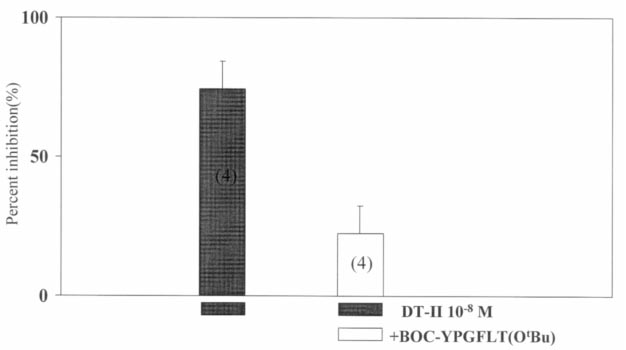 Figure 4. The reversal of the inhibitory action of deltorphin
Figure 4. The reversal of the inhibitory action of deltorphin