Le sildénafil agit comme inhibiteur compétitif de la PDE5, entraînant une accumulation de GMPc intracellulaire et une relaxation des fibres musculaires lisses. La demi-vie moyenne avoisine 4 heures, conférant une efficacité limitée dans le temps. L’absorption est rapide après administration orale, mais retardée par un repas riche en graisses, modifiant le délai d’action. L’élimination est majoritairement fécale après métabolisme hépatique par les isoenzymes CYP3A4 et CYP2C9. Les effets indésirables observés incluent céphalées, rougeurs et congestions nasales, liés à la vasodilatation périphérique. Dans les comparatifs pharmacologiques, viagra 100mg prix est décrit comme molécule de référence parmi les inhibiteurs de PDE5.
9903064
Allergen-induced Synthesis of F2-Isoprostanes in Atopic Asthmatics Evidence for Oxidant Stress RYSZARD DWORSKI, JOHN J. MURRAY, L. JACKSON ROBERTS II, JOHN A. OATES, JASON D. MORROW, LAURA FISHER, and JAMES R. SHELLER
Center for Lung Research and Department of Medicine, Vanderbilt University School of Medicine, Nashville, Tenessee
It is thought that reactive oxygen species (ROS) participate in the inflammation which characterizes asthma, but the evidence supporting this contention is incomplete. F2-isoprostanes (F2-IsoPs) are arachidonate products formed on membrane phospholipids by the action of ROS and thereby repre- sent a quantitative measure of oxidant stress in vivo. Using a mass spectrometric assay we measured urinary release of F2-IsoPs in 11 patients with mild atopic asthma after inhaled allergen challenge. The excretion of F 2-IsoPs increased at 2 h after allergen (1.5 0.2 versus 2.6 0.3 ng/mg creatinine) and remained significantly elevated in all urine collections for the 8-h period of the study (analysis of variance [ANOVA]). The measured compounds were of noncyclooxygenase origin because neither aspirin nor indomethacin given before challenge suppressed them. Urinary F2-IsoPs remained un- changed after inhaled methacholine challenge. In nine atopic asthmatics, F2-IsoPs were quantified in bronchoalveolar lavage fluid (BALF) at baseline values and in a separate segment 24 h after allergen instillation. F 2-IsoPs were elevated late in the BALF (0.9 0.2 versus 11.4 3.0 pg /ml, baseline ver- sus allergen, respectively, p ϭ 0.007). The increase was inhibited by pretreatment of the subjects with inhaled corticosteroids. These findings provide a new evidence for a role for ROS and lipid peroxida- tion in allergen-induced airway inflammation. Dworski R, Murray JJ, Roberts LJ, II, Oates JA, Mor- row JD, Fisher L, Sheller JR. Allergen-induced sythesis of F2-isoprostanes in atopic asthmat- ics: evidence for oxident stress. AM J RESPIR CRIT CARE MED 1999;160:1947–1951.
Asthma is a chronic inflammatory disease of the airways of
the reaction of nitric oxide (.NO) with superoxide (OϪ2) gives
unknown origin which is increasing in prevalence. A profound
peroxynitrite (ONOO)/peroxynitrous acid (ONOOH), a pow-
inflammation is a characteristic feature of fatal asthma. How-
erful indiscriminate oxidant and nitrating agent. Recently, it
ever, recent studies have clearly shown that allergen-mediated
has been shown that peroxynitrite is produced in the asth-
inflammation exists even in patients with mild disease (1). Re-
matic airway (9). The finding is not surprising because in-
active oxygen species (ROS) are likely to participate in asthma.
creased amounts of both .NO and O are generated in asthma
There is a favorable biochemical environment for free radical
(10). In addition to cells, there are several environmental sources
mediated reactions in the asthmatic airways: (1) the major in-
of oxidants, e.g., ozone, a major pollutant, is a potent nonradi-
flammatory cells involved in asthmatic inflammation such as
cal oxidant known to generate free radicals in the airways in
macrophages and eosinophils produce ROS when activated
vivo (11). Nevertheless, because of substantial difficulties in
with different stimuli, and, moreover, cells from asthmatics
the quantitative measurement of oxidant stress in vivo much
possess an increased capability to generate free radicals com-
of the evidence for the activity of ROS in asthmatic inflamma-
pared with normal cells (2–4); (2) the oxidant “burst” in the
tion is indirect or circumstantial. Thus, measurement of in-
cells can be stimulated by some asthma mediators (5–7); and
creases in .NO and H2O2 in exhaled gas has been taken as an
(3) antioxidant mechanisms are disturbed in asthmatics (8).
indication of ROS production, but evidence showing that this
Although the commonly generated ROS superoxide radical
results in oxidative consequences in the airway is lacking.
(OϪ2) and hydrogen peroxide (H2O2) per se can oxidize biolog-
F2-Isoprostanes (F2-IsoPs) are recently discovered stable
ical substrates, the damaging effect is greater when they react
prostaglandin-like compounds that are primarily synthesized
with one another or with other reactive species. For example,
by free radical catalyzed peroxidation of arachidonic acid in-dependent of the cyclooxygenase (COX) enzyme. Four F2-IsoP regioisomers can be formed and each of them can consti-
(Received in original form March 11, 1999 and in revised form June 7, 1999 )
tute eight racemic diastereomers giving a total of 64 different
Supported by NIH Grants GM 15431, GM 42056, DK 48831, CA 77839, DK
compounds (12, 13). Unlike prostaglandins, isoprostanes re-
main in the cell membrane phospholipids until hydrolyzed by
Correspondence and requests for reprints should be addressed to Ryszard Dwor-
specific phospholipases (14). E- and D-ring IsoPs, isothrom-
ski, M.D., Center for Lung Research, Vanderbilt University School of Medicine, T-
boxanes, and isoleukotrienes have also been reported. Re-
1217 Medical Center North, Nashville, TN 37232-2650. E-mail: ryszard.dworski@
Am J Respir Crit Care Med Vol 160. pp 1947–1951, 1999
to assess oxidant status in vitro and in vivo in animals and hu-
Internet address: www.atsjournals.org
mans. F2-IsoPs are elevated in a number of human vascular
AMERICAN JOURNAL OF RESPIRATORY AND CRITICAL CARE MEDICINE
and inflammatory disorders which have been thought to be as-
monary function measurements were made 3 min after each dose un-
sociated with an oxidant stress such as coronary reperfusion,
til the FEV1 had decreased by 20% from baseline values. All proto-
atherosclerosis, hypercholesterolemia, hepatorenal syndrome,
cols were approved by the Vanderbilt University Committee for the
liver cirrhosis, diabetes mellitus, scleroderma, and in smokers
Protection of Human Subjects. All volunteers signed written consentforms before proceeding with the study.
(13). In the human lung, increased formation of F2-IsoPs hasbeen demonstrated in bronchoalveolar lavage fluid (BALF)
Analytical Methods
after exposure to ozone (15), in patients with interstitial pul-
F -IsoPs in urine and BALF were measured by stable isotope dilution
monary fibrosis (16), and in chronic obstructive pulmonary
assay that used gas chromatography/negative-ion chemical ionization
mass spectrometry (GC-NICI-MS) (20). In brief, the specimens were
In the present study, we demonstrate that inhaled allergen
acidified to pH 3 with 1 M HCl and deuterated internal standard
([ H4]8-epi-prostaglandin F2 alpha) was added. Then the samples were
atopic asthmatics. In addition, F2-IsoPs are produced in BALF
extracted on C18 Sep-Pak columns (Waters Chromatography Division,
24 h after segmental allergen challenge, and the increase can
Millipore, Milford, MA) and converted to pentafluorobenzyl (PFB)
be abolished by pretreatment of the volunteers with inhaled
esters by treatment with a mixture of 10% pentafluorobenzyl bromide
corticosteroids. Thus we provide direct evidence that oxidant
and 10% N,N-diisopropylethylamine in acetonitrile (Aldrich Chemical
injury occurs in the setting of allergic inflammation.
Division, Milwaukee, WI). After evaporation of the reagents underN2, the residue was subjected to thin layer chromatography (What-man, Inc., Clifton, NJ). The plates were scraped according to standard
and eluted from the silica with ethyl acetate. The samples were dried
Subjects
under N2 and converted to trimethylsilyl ether derivatives by addingN,O-bis(trimethylsilyl)trifluoroacetamide (BSTFA) in pyridine (Ald-
Eleven patients with mild atopic asthma, 21 to 44 yr of age, underwent
rich Chemical Division). The analysis of F2-IsoPs was performed us-
inhaled allergen challenge. Bronchoscopy with bronchoalveolar la-
ing a Nermag R10-10C (Fairfield, NJ) or Hewlett-Packard 5982A
vage (BAL) was performed in nine mild allergic asthmatics 24 to 46 yr
mass spectrometer (Palo Alto, CA) and a 15-m DB1701 fused silica
of age. None of the subjects was using corticosteroids, disodium cromo-
capillary column (J&W Scientific, Folsom, CA). Ions were monitored
glycate, or antileukotriene agents. The patients were asked to discon-
at a mass-to-charge ratio (m/z) of 569 for endogenous F2-IsoPs and at
tinue theophylline and antihistamines for 48 h, and inhaled short-acting
-agonist for at least 12 h before allergen challenges. All volunteers
PGD-M (9␣, 11-dihydroxy-15-oxo-2,3,18,19-tetranorprost-5-ene-
were skin-test positive to at least one allergen.
1,20-dioic acid), the major urinary metabolite of prostaglandin D2-PGD2, was quantified by GC-NICI-MS as previously described (21). Study Design
Briefly, to 1 ml of urine [18O4] PGD-M internal standard was added.
A screening allergen inhalation challenge to determine the dose of al-
Then the sample was acidified to pH 3 with HCl and left to stand at
room temperature for 30 min to allow quantitative cyclization of the
dose), and the measurement of spirometry were carried out as previ-
lower side chain to a hemiketal lactone. After extraction with a C18
ously described (18). Antigens used included grass, dust mite, and cat
Sep-Pak, methylation of the upper carboxyl, and methoxymation of
(Bayer, Spokane, WA). At least 2 wk after the screening challenge,
the keto group at C-15, borate buffer (pH 9.1) was added and neutral
subjects underwent provocation with the threshold dose of allergen.
lipids were extracted with ethyl acetate. The aqueous layer was then
Pulmonary function was measured at baseline, then every 15 min for
acidified to pH 3 with HCl, and PGD-M was extracted with methylene
the first hour after allergen inhalation, and then hourly for 8 h after in-
chloride. The lower carboxyl was then converted to a pentafluoroben-
halation challenge. Urine for analysis of F
zyl ester, and the partially derivatized PGD-M was purified on thin
challenge and then every 2 h for the 8 h of the study. On a separate oc-
layer chromatography. After conversion to a trimethylsilyl ether de-
casion, inhaled allergen challenge and the collection of urine samples
rivative, quantification of PGD-M was accomplished by selected ion
were reproduced in six volunteers after pretreatment with oral aspirin
monitoring: mass-to-charge ratio was 514 for endogenous PGD-M and
(three doses of 900 mg the day before and 900 mg on the morning of
the study) or indomethacin (50 mg in three daily doses for 2 d and 50mg on the morning of the study and at noon). In another four volun-
Statistical Evaluation
teers urine samples were collected at baseline and in 2-h intervals for
Kolmogorow-Smirnov testing showed normal distribution of the data.
8 h after a challenge with inhaled methacholine causing a decrease in
Consequently, urinary eicosanoids were analyzed using repeated mea-
sures analysis of variance (ANOVA) and Student-Newman-Keuls
The bronchoscopy study was conducted according to the protocol
multiple comparisons test. Analysis of F2-IsoPs in BALF was per-
described elsewhere (19). Nine subjects were randomly assigned to
formed using the Student’s t test. Significance was accepted when the
receive either inhaled beclomethasone (6 puffs of 42 g each 1 h be-
fore antigen instillation, and then the same dose 12 and 24 h later) oridentical placebo in a double-blind, crossover fashion. After topical
lidocaine anesthesia, a fiberoptic bronchoscope was inserted into theairways, and a control BAL was performed in either a lingula or right
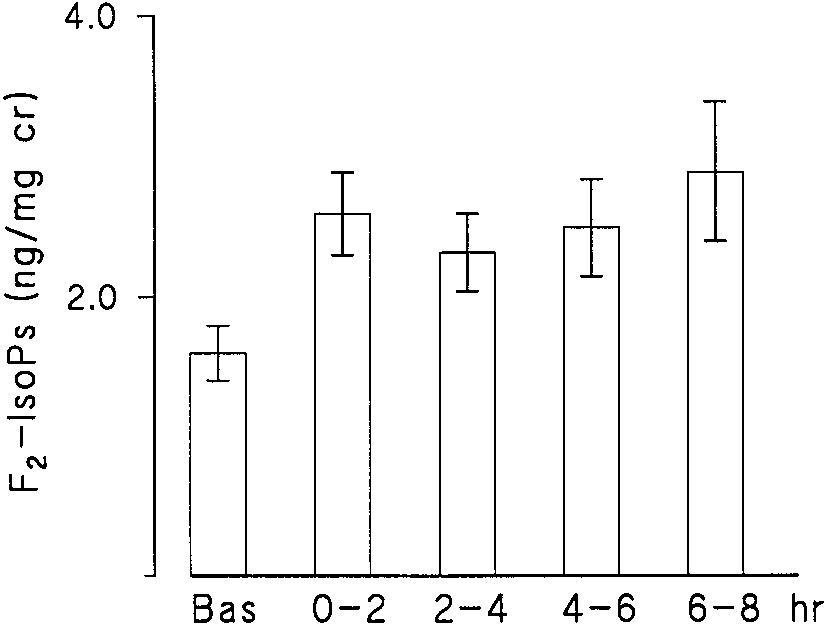
In all volunteers inhalation of the allergen provoked a 20% or
middle lobe using 50-ml aliquots of warmed normal saline. In the op-
greater decrease in FEV1 from the baseline. The excretion of
posite segment, allergen to which the volunteer was skin prick test pos-
F2-IsoPs was significantly increased at 2 h after allergen inha-
itive was instilled (5 ml of a solution at 10 allergy units [AU] or 1:10,000).
lation and remained elevated in all urine collections for the
Drug was continued and 24 h later the antigen-challenged segment
8-h time period of the study (Figure 1). Six patients developed
was similarly lavaged. After a washout period of 3 wk or greater, pa-
a late response, which was defined as a decrease in FEV of 20%
tients were crossed over to the other arm of the study. All patients tol-
erated the study without incident. The fluid was filtered through loose
or more from the baseline at 4 to 8 h after allergen inhalation.
gauze, centrifuged, and stored at Ϫ70Њ C until the analysis was done.
The concentrations of F2-IsoPs in these patients were not in-creased compared with subjects with only an early response. Methacholine Challenge
To demonstrate that the F2-IsoPs appearing in the urine af-
ter allergen challenge were not the COX products of arachi-
Volunteers had measurements of FEV1 in duplicate followed by inha-lation of doubling concentrations of methacholine every 5 min start-
donic acid, oral aspirin or indomethacin was administered in
ing at a concentration 0.075 mg/ml to a maximum of 20.0 mg/ml via
six volunteers before allergen inhalation challenge. COX inhi-
DeVilbiss 646 nebulizer coupled to a Rosenthal-French dosimeter
bition was documented by the measurement of the urinary lev-
(Laboratory of Applied Immunology, Baltimore, MD). Repeat pul-
els of PGD-M, a metabolite of PGD2, the major COX product
Dworski, Murray, Roberts, et al.: F2-Isoprostanes in Allergen-induced Asthma
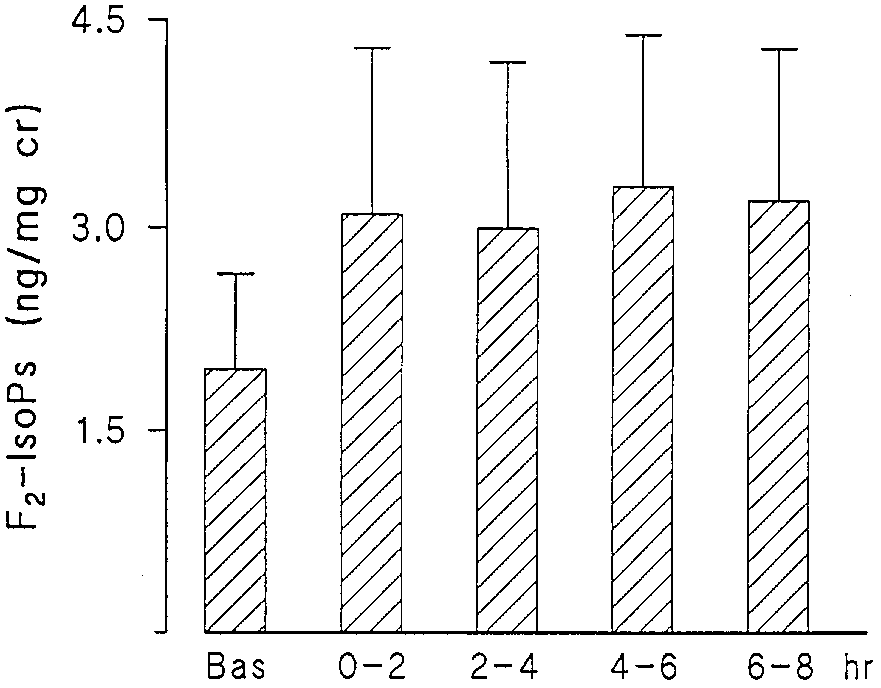
Figure 1. Urinary concentrations of F2-IsoPs in ng/mg creatinine versus time in 11 atopic asthmatics at baseline and after allergen Figure 3. Urinary F
2-IsoPs in six atopic asthmatics at baseline and
after allergen inhalation challenge after a COX inhibitor. Height of
riod is shown as Ϯ SEM. The differences among columns were sig-
the bars indicates mean Ϯ SEM. The differences among columns
of mast cells. As shown in Figure 2, aspirin or indomethacin
erated during allergen-induced reaction in atopic asthmatics.
was effective in blocking the allergen-stimulated increase in
Increased concentrations of isoprostanes, determined by the
PGD-M. In contrast, the levels of F2-IsoPs were not sup-
sensitive and specific GC-NICI-MS method, were found in the
pressed, confirming a non-COX origin of the measured com-
urine after inhaled allergen challenge and in BALF in re-
pounds (Figure 3). To ensure that the production of F2-IsoPs
sponse to segmental deposition of the antigen. The formation
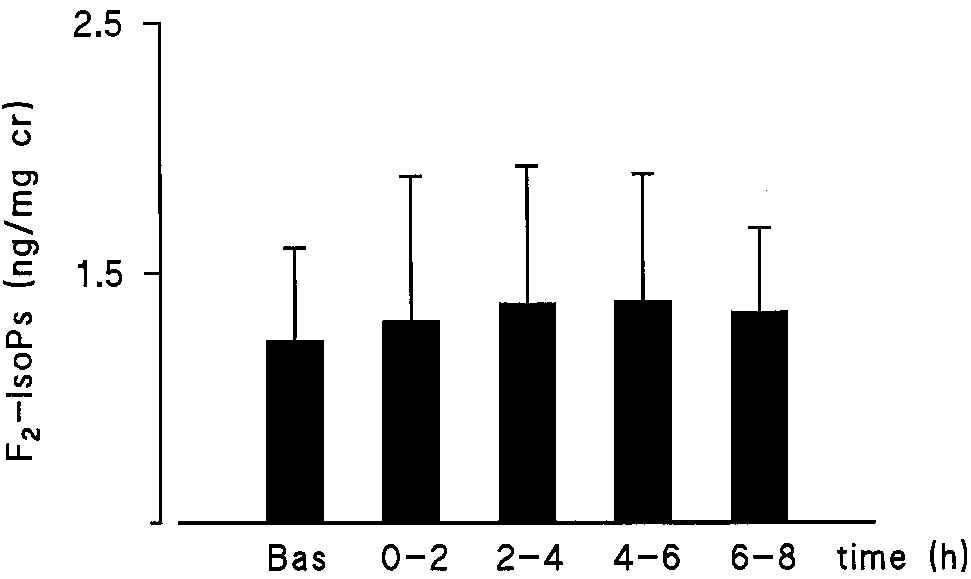
was specific to the inhaled allergen, four volunteers under-
of F -IsoPs appears to be a specific response to allergen be-
went a challenge with inhaled methacholine causing a 20% or
cause the nonspecific bronchoconstrictor, methacholine, did
greater decrease in FEV1. No alterations in urinary concentra-
not cause an increase in F -IsoPs. The measured compounds
2-IsoPs were generated by methacholine (Figure 4).
were non-COX products because they were not abrogated by
Subsequently, F2-IsoPs were measured in BALF from nine
pretreatment of the subjects with either aspirin or indometha-
patients with mild atopic asthma at baseline, and from a sepa-
cin. The inhibition of the COX enzyme in the subjects proved
rate lung segment 24 h after allergen instillation. There was a
to be adequate because the synthesis of PGD assessed by the
measurement of its major urinary metabolite, PGD-M, was
allergen challenge. The response was inhibited by pretreat-
efficiently blocked by either aspirin or indomethacin. COX-
ment of the volunteers with inhaled corticosteroids (Figure 5).
dependent formation of F2-IsoPs has been reported (22), butCOX enzymatic activity does not appear to be the source of
DISCUSSION
the urinary F2-IsoPs generated as a consequence of allergic
airway stimulation. It is important to emphasize that the fam-
lyzed peroxidation lipid products of arachidonic acid, are gen-
ily of F2-IsoPs contains 64 different compounds which havebeen identified both in vitro and in vivo (13) and that ourmethodology does not characterize the whole profile of F2-ring isoprostanes produced during allergen-evoked inflamma-tion in asthmatics; thus, the total amount of isoprostanes gen-
Figure 2. Urinary concentrations of PGD-M in ng/mg creatinine in six atopic asthmatics at baseline and after allergen provocation af- ter placebo (open bars) or a COX inhibitor (hatched bars). Mean Figure 4. Urinary F2-IsoPs in four atopic asthmatics at baseline and
amount of PGD-M for each time period is shown as Ϯ SEM. Aspi-
after methacholine inhalation challenge. Height of the bars indi-
rin or indomethacin significantly blocked the allergen-stimulated
cates mean Ϯ SEM. The release of F2-IsoPs was unchanged by
methacholine inhalation causing 20% or greater decrease in FEV1.
AMERICAN JOURNAL OF RESPIRATORY AND CRITICAL CARE MEDICINE
can be interpreted as new evidence for an increased oxidantstress in allergen-induced asthmatic reaction. Our finding co-incides with the results of earlier studies demonstrating theaugmented activity of ROS in asthma (3, 4, 11, 24–26). The in-hibition of allergen-provoked formation of F2-IsoPs in BALFby inhaled beclomethasone suggests that corticosteroids,which are known to inhibit the late allergic inflammation, mayact in part by restraining oxidant stress. This effect of corticos-teroids may have been caused by a reduction in the numberand activation of cells producing free radicals. Indeed, in asimilar experimental model of the late-phase reaction to localallergen challenge, inhaled corticosteroids diminished the in-flux of eosinophills and reduced the production of inflamma-tory mediators such as leukotriene B4 (27).
It is unknown if F2-IsoPs play a role as a pathophysiological
factor in allergen-provoked asthma. Isoprostanes are biologi-cally active compounds. Some of the known effects of F2-IsoPscould be relevant to the pathophysiology of the lung. For ex-ample, one of the F2-IsoPs, 8-epi-PGF2␣, constricts animal and
human airways in vitro (28) and causes airflow obstruction andairway plasma exudation in guinea pig in vivo (29). 8-epi-PGF2␣ is also a potent vasoconstrictor of the pulmonary artery
Figure 5. F2-IsoPs in pg/ml BAL fluid from nine atopic asthmatics
in rabbits and rats (13, 30). Therefore, although no analogous
at baseline and after allergen instillation on placebo (Pl) and 24 h
data exist in humans, F2-IsoPs could be viewed not only as
after pretreatment with inhaled corticosteroids (CS). Height of the
markers but also as possible mediators of oxidant stress injury
bars indicates mean Ϯ SEM. Allergen challenge provoked a signifi-
in vivo. The mechanism of the action of F2-IsoPs is unclear.
cant increase in F2-IsoPs on placebo (open bars) (p ϭ 0.007). The in-
The involvement of the thromboxane receptor as well as a
crease was suppressed by pretreatment with inhaled beclometha-
unique isoprostane receptor has been proposed, though the
sone (hatched bars) (p ϭ 0.13).
latter hypothesis has not been validated by any experimentaldata (13). While most of the studies on the biological functionsof F2-IsoPs have been performed with 8-epi-PGF2␣, it is likely
erated is unknown but must be greater than the amount we
that other compounds are also bioactive (31). Finally, one can
not exclude the possibility that other biologically active iso-
The rise of the concentrations of F2-IsoPs in BALF 24 h af-
prostanes (for example, isoprostanes of the E series and isole-
ter segmental allergen challenge shows that isoprostanes are
ukotrienes [13] and even distinct classes of lipid peroxidation
generated directly in the airways of asthmatic subjects in re-
products) could also be formed in response to allergen in asth-
sponse to allergen. On the other hand, we did not find consis-
matics. This notion leads to another general question: could
tent release of F2-IsoPs into BALF 4 min after allergen in-
ROS-generated oxidized lipids play a pathophysiological role
stillation (data not shown). The lack of F2-IsoPs early in an
in allergic inflammation? Recent experimental studies sug-
allergic reaction could result from a different time course of
gest that modest oxidation of membrane lipids may stimulate
ROS formation as opposed to mediators such as tryptase and
the expression of selected genes and alter several cellular re-
the leukotrienes. It is also known that there is a significant de-
sponses (32–35). From that perspective, investigation of ROS-
lay from the time of initial lipid peroxidation on the cell mem-
mediated lipid peroxidation might be an exciting and impor-
brane to the appearance of free isoprostanes (14). This pre-
tant avenue for future studies on the pathogenesis of allergic
sumably results from the time necessary for phospholipase
activation and subsequent hydrolysis of isoprostanes.
In summary, this study demonstrates that inhaled allergen
The quantitative assessment of oxidant stress in pathophys-
challenge causes the release of F2-IsoPs into the urine of
iological processes, particularly in vivo, has been associated
atopic asthmatics. F2-IsoPs are also increased in BALF late af-
with major difficulties due to the deficiency of reliable meth-
ter segmental allergen challenge, and this increase can be re-
ods. The analysis of lipid peroxidation in human body fluids
duced by pretreatment with inhaled corticosteroids. The patho-
and tissues based on diene-conjugate and thiobarbituric acid
physiological role of lipid peroxidation in the pathogenesis of
(TBA) assays has been frequently used for that purpose; how-
asthmatic inflammation provoked by allergen challenge is cur-
ever, both methods are characterized by a low sensitivity and
specificity and, furthermore, can produce confusing artifacts(23). Other traditional approaches, such as an analysis of ex-
Acknowledgment : The authors thank Brendie Keane, R.N., and John Hols-
pired breath condensate hydrogen peroxide and exhaled pen-
inger for their assistance with this study, and Tamara Lasakow for editorialhelp in preparing the manuscript.
tane levels or a measurement of substrate oxidizability or spintrapping of free radical adducts ex vivo suffer from similar lim-itations. Isoprostanes are stable free radical–catalyzed prod-
References
ucts of arachidonic acid. There has been a growing number of
1. Beasley, R., W. R. Roche, J. A. Roberts, and S. T. Holgate. 1989. Cellu-
studies indicating that a specific quantification of F2-IsoPs in
lar events in the bronchi in mild asthma and after bronchial provoca-
biological samples is a sensitive and a reliable noninvasive
tion. Am. Rev. Respir. Dis. 139:806–817.
method allowing assessment of ROS-caused lipid peroxida-
2. Sedgwick, J. B., K. M. Geiger, and W. W. Busse. 1990. Superoxide gen-
eration by hypodense eosinophils from patients with asthma. Am.
tion and oxidant stress in vivo (13). Therefore, the elevated
Rev. Respir. Dis. 142:120–125.
concentrations of F2-IsoPs in the urine and BAL samples from
3. Calhoun, W. J., H. E. Reed, D. R. Moest, and C. A. Stevens. 1992. En-
asthmatic subjects after allergen challenge shown in our study
hanced superoxide production by alveolar macrophages and air-space
Dworski, Murray, Roberts, et al.: F2-Isoprostanes in Allergen-induced Asthma
cells, airway inflammation, and alveolar macrophage density changes
Sheller, J. E. Fish, and S. P. Peters. 1966. A controlled trial of the ef-
after segmental antigen bronchoprovocation in allergic subjects. Am.
fect of the 5-lipoxygenase inhibitor, zileuton, on lung inflammation
Rev. Respir. Dis. 145:317–325.
produced by segmental antigen challenge. J. Allergy Clin. Immunol.
4. Sanders, S. P., J. L. Zweier, S. J. Harrison, M. A. Trush, S. J. Rembish,
and M. C. Liu. 1995. Spontaneous oxygen radical production at sites
20. Morrow, J. D., and L. J. Roberts, II. 1998. Mass spectrometric quantifi-
of antigen challenge in allergic subjects. Am. J. Respir. Crit. Care Med.
cation of F2-isoprostanes in biological fluids and tissues as measure of
oxidant stress. Methods Enzym. 300:3–12.
5. Chihara, J., N. Hayashi, T. Kakazu, T. Yamamoto, D. Kurachi, and S.
21. Morrow, J. D., C. Prakash, J. A. Awad, T. A. Duckwort, W. E. Zackert,
Nakajima. 1996. RANTES augments radical oxygen products from eo-
I. A. Blair, J. A. Oates, and L. J. Roberts, II. 1991. Quantification of
sinophils. Intern. Arch. Allergy Immunol. 104(Suppl.):52–53.
the major urinary metabolite of prostaglandin D2 by a stable isotope
6. Tenscher, K., B. Metzner, E. Schopf, J. Norgauer, and W. Czech. 1996.
dilution mass spectrometric assay. Anal. Biochem. 193:142–148.
Recombinant human eotaxin induces oxygen radical production,
22. Pratico, D., J. A. Lawson, and G. A. FitzGerald. 1995. Cyclooxygenase-
Ca(2ϩ)-mobilization, actin reorganization, and CD11b upregulation
dependent formation of the isoprostane, 8-epi-prostaglandin F2 alpha.
in human eosinophils via a pertussis toxin-sensitive heterotrimeric
J. Biol. Chem. 270:9800–9808.
guanine nucleotide-binding protein. Blood 88:3195–3199.
23. Halliwell, B., and S. Chirico. 1993. Lipid peroxidation: its mechanism, mea-
7. Bruijnzeel, P. L. B., L. Koenderman, P. T. M. Kok, M. L. Hamelink, and
surement, and significance. Am. J. Clin. Nutr. 57(Suppl.):715S–725S.
J. L. Verhagen. 1986. Platelet activating factor (PAF-acether) induced
24. Dohlman, A. W., H. R. Black, and J. A. Royall. 1993. Expired breath hy-
leukotriene C4 formation and luminol dependent chemiluminescence
drogen peroxide is a marker of acute airway inflammation in pediatric
of human eosinophils. Pharm. Res. Comm. 18:61–69.
patients. Am. Rev. Respir. Dis. 148:955–960.
8. De Raeve, H. R., F. B. J. M. Thunnissen, F. T. Kaneko, F. H. Guo, M.
25. Jarjour, N. N., W. W. Busse, and W. J. Calhoun. 1992. Enhanced produc-
Lewis, M. S. Kavuru, M. Secic, M. J. Thomassen, and S. C. Erzurum.
tion of oxygen radicals in nocturnal asthma. Am. Rev. Respir. Dis. 146:
1997. Decreased Cu,Zn-SOD activity in asthmatic airway epithelium:
correction by inhaled corticosteroid in vivo. Am. J. Physiol. 272:L148–
26. Antczak, A., D. Nowak, D., B. Shariati, M. Król, G. Piasecka, and Z.
Kurmanowska. 1997. Increased hydrogen peroxide and thiobarbituric
9. Saleh, D., P. Ernst, S. Lim, P. J. Barnes, and A. Giaid. 1998. Increased
acid–reactive products in expired breath condensate of asthmatic pa-
formation of the potent oxidant peroxynitrite in the airways of asth-
tients. Eur. Respir. J. 10:1235–1241.
matic patients is associated with induction of nitric oxide synthase: ef-
27. Murray, J. J., D. D. Hagaman, R. Dworski, B. Keane, and J. R. Sheller.
fect of inhaled glucocorticoid. FASEB J. 12:929–937.
1988. Inhibition by salmeterol and beclomethasone of late phase re-
10. Kharitonov, S. A., D. Yates, R. A. Robbins, R. Logan-Sinclair, E. A.
sponse to segmental antigen challenge in asthmatics (abstract). Am. J.
Shinebourne, and P. J. Barnes. 1994. Increased nitric oxide in exhaled
Respir. Crit. Care Med. 157:A872.
air of asthmatic patients. Lancet 343:133–135.
28. Kawikova, I., P. J. Barnes, T. Takahashi, S.Tadjkarimi, M. H. Yacoub,
11. Mustafa, M. G. 1990. Biochemial basis of ozone toxicity. Free Rad. Biol.
and M. G. Belvisi. 1996. 8-epi-PGF2␣, a novel noncyclooxygenase-
derived prostaglandin, constricts airways in vitro.Am. J. Respir. Crit.
12. Morrow, J. D., K. E. Hill, R. F. Burk, T. M. Nammour, K. F. Badr, and
L. J. Roberts, II. 1990. A series of prostaglandin F2-like compounds
29. Okazawa, A., I. Kawikova, Z.-H. Cui, B-E. Skoogh, and J. Lötvall. 1997.
are produced in vivo in humans by a non-cyclooxygenase, free radi-
8-epi-PGF2␣ induces airflow obstruction and airway plasma exudation
cal–catalyzed mechanism. Proc. Natl. Acad. Sci. U.S.A. 87:9383–9387. in vivo.Am. J. Respir. Crit. Care Med. 155:436–441.
13. Morrow, J. D., and L. J. Roberts, II. 1997. The isoprostanes: unique bio-
30. Banerjee, M., K. Kang, J. D. Morrow, L. J. Roberts, II, and J. H. New-
active products of lipid peroxidation. Prog. Lipid Res. 36:1–21.
man. 1992. Effect of a novel prostaglandin, 8-epi-PGF2␣ in rabbit lung
14. Morrow, J. D., J. A. Awad, H. J. Boss, I. A. Blair, and L. J. Roberts, II.
in situ. Am. J. Physiol. 263:H660–H663.
1992. Non-cyclooxygenase-derived prostanoids (F2-isoprostanes) are
31. Kunapuli, P., J. A. Lawson, J. A. Rokach, J. L. Meinkoth, and G. A.
formed in situ on phospholipids. Proc. Natl. Acad. Sci. U.S.A. 89:
FitzGerald. 1998. Prostaglandin F2␣ (PGF2␣) and the isoprostane, 8,12-
iso-isoprostane F2␣-III, induce cardiomyocyte hypertrophy. J. Biol.
15. Hazbun, M. E., R. Hamilton, A. Holian, and W. L. Eschenbacher. 1993.
Ozone-induced increases in substance P and 8-epi-prostaglandin F2␣ in
32. Suzuki, Y. J., H. F. Forman, and A. Sevanian. 1997. Oxidants as stimula-
the airways of human subjects. Am. J. Respir. Cell Mol. Biol. 9:568–572.
tors of signal transduction. J. Free Rad. Biol. Med. 22:269–285.
16. Montuschi, P., G. Ciabattoni, P. Paredi, P. Pantelidis, R. M. Du Bois,
33. Rashba-Step, J. A., A. Tatoyan, R. Duncan, A. Ann, T. R. Puspa-Rehka,
S. A. Kharitonov, and P. J. Barnes. 1998. 8-Isoprostane as a biomarker
and A. Sevanian. 1997. Phospholipid peroxidation induces cytosolic
of oxidative stress in interstitial lung diseases. Am. J. Respir. Crit. Care
phospholipase A2 activity: membrane effects versus enzyme phospho-
rylation. Arch. Biochem. Biophys. 343:44–54.
17. Pratico, D., S. Basili, M. Vieri, C. Cordova, F. Violi, and G. A. FitzGer-
34. Leonardduzzi, G., A. Scavazza, F. Biasi, E. Chiarpotto, S. Camandola, S.
ald. 1998. Chronic obstructive pulmonary disease is associated with an
Vogl, R. Dargel, and G. Poli. 1997. The lipid peroxidation end product
increase in urinary levels of isoprostane F2␣-III, an index of oxidant
4-hydroxy-2-3-nonenal up-regulates transforming growth factor 1 ex-
stress. Am. J. Respir. Crit. Care Med. 158:1709–1714.
pression in the macrophage lineage: a link between oxidative injury
18. Sladek, K., R. Dworski, G. A. FitzGerald, K. L. Buitkus, F. J. Block,
and fibrosclerosis. FASEB J. 11:851–857.
S. R. Marney, Jr., and J. R. Sheller. 1990. Allergen-stimulated release
35. Kramer, K., B. Rademaker, W.H. Rozendal, H. Timmerman, and A.
of thromboxane A2 and leukotriene E4 in humans. Am. Rev. Respir.
Bast. 1986. Influence of lipid peroxidation on beta-adrenoceptors.
19. Kane, G. C., M. Pollice, M. Kim, C. J. Cohn, R. Dworski, J. J. Murray, J.
DIE FILES DÜRFEN NUR FÜR DEN EIGENEN GEBRAUCH BENUTZT WERDEN. DAS COPYRIGHT LIEGT BEIM JEWEILIGEN AUTOR. COMER: WICHTIGE NAMEN UND MODELLE, KAPITEL 1-10 Somatogene Sichtweise: Emil Kraeplin (vs. psychogene Sichtweise u.a. mit Hypnose, Josef Breuer)) Deinstitutionalisierung und gemeindepsychiatrischer Ansatz Gehirn, synaptischer Spalt, Neurotransmitter Pawlow, Watson, Thornd
Annex 6.1 QS Active Agent Catalogue for Poultry The subsequent table contains a complete list of the active substances in veterinary medicines which are approved in Germany for application to poultry. In consideration of the various domestic regulations regarding the waiting period and duration, the following applies: 1. Legally binding is invariably the waiting period indicated on the formu
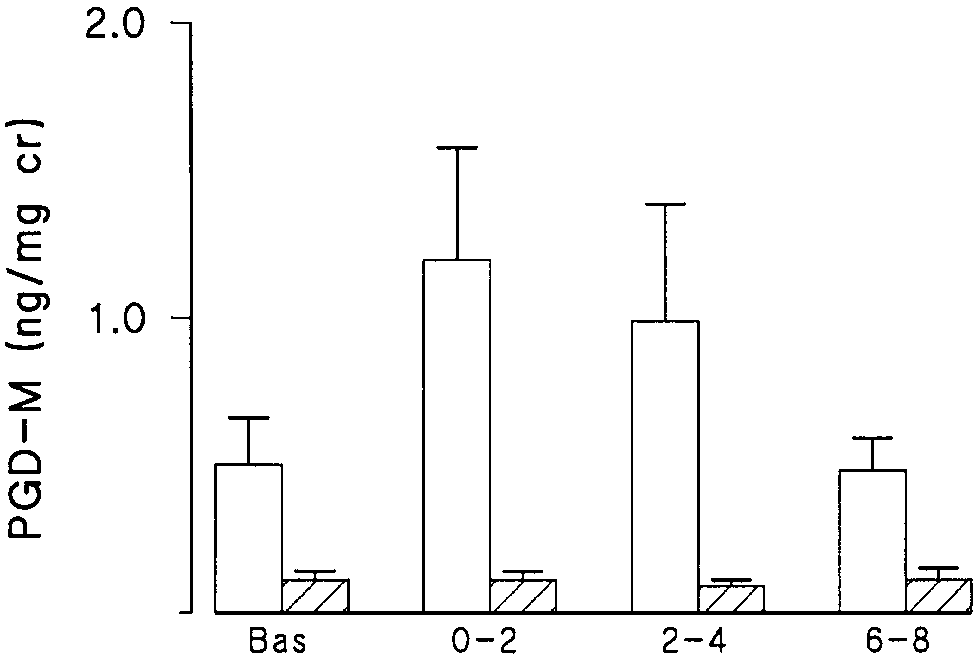
 Dworski, Murray, Roberts, et al.: F2-Isoprostanes in Allergen-induced Asthma
Figure 1. Urinary concentrations of F2-IsoPs in ng/mg creatinine
Dworski, Murray, Roberts, et al.: F2-Isoprostanes in Allergen-induced Asthma
Figure 1. Urinary concentrations of F2-IsoPs in ng/mg creatinine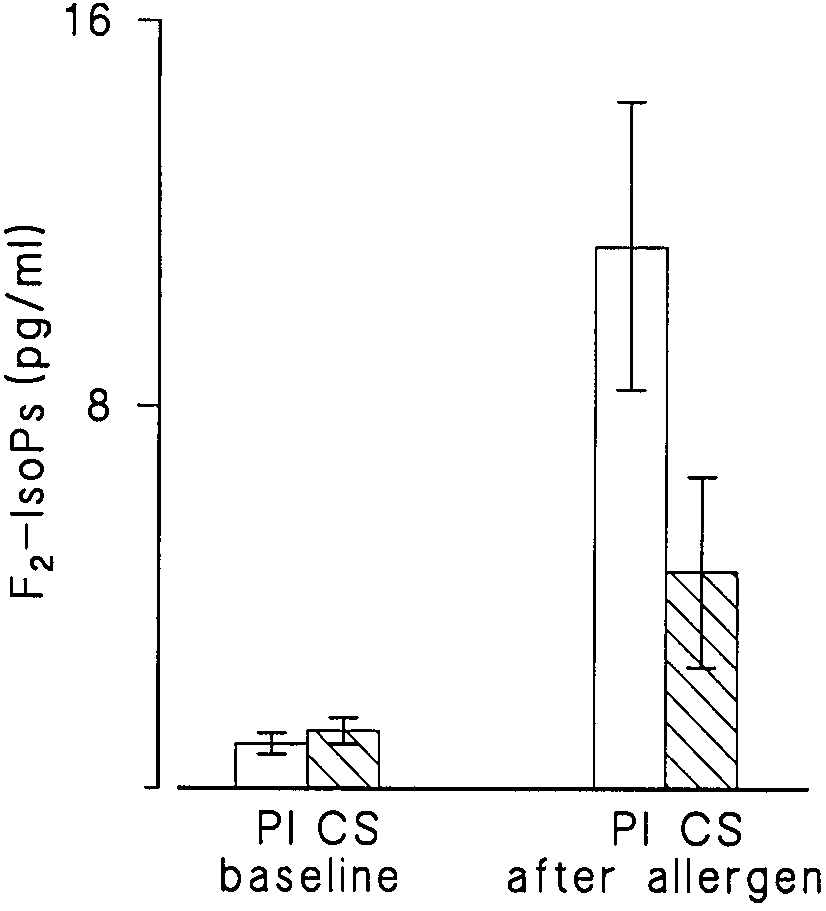 AMERICAN JOURNAL OF RESPIRATORY AND CRITICAL CARE MEDICINE
can be interpreted as new evidence for an increased oxidantstress in allergen-induced asthmatic reaction. Our finding co-incides with the results of earlier studies demonstrating theaugmented activity of ROS in asthma (3, 4, 11, 24–26). The in-hibition of allergen-provoked formation of F2-IsoPs in BALFby inhaled beclomethasone suggests that corticosteroids,which are known to inhibit the late allergic inflammation, mayact in part by restraining oxidant stress. This effect of corticos-teroids may have been caused by a reduction in the numberand activation of cells producing free radicals. Indeed, in asimilar experimental model of the late-phase reaction to localallergen challenge, inhaled corticosteroids diminished the in-flux of eosinophills and reduced the production of inflamma-tory mediators such as leukotriene B4 (27).
AMERICAN JOURNAL OF RESPIRATORY AND CRITICAL CARE MEDICINE
can be interpreted as new evidence for an increased oxidantstress in allergen-induced asthmatic reaction. Our finding co-incides with the results of earlier studies demonstrating theaugmented activity of ROS in asthma (3, 4, 11, 24–26). The in-hibition of allergen-provoked formation of F2-IsoPs in BALFby inhaled beclomethasone suggests that corticosteroids,which are known to inhibit the late allergic inflammation, mayact in part by restraining oxidant stress. This effect of corticos-teroids may have been caused by a reduction in the numberand activation of cells producing free radicals. Indeed, in asimilar experimental model of the late-phase reaction to localallergen challenge, inhaled corticosteroids diminished the in-flux of eosinophills and reduced the production of inflamma-tory mediators such as leukotriene B4 (27).