Le sildénafil agit comme inhibiteur compétitif de la PDE5, entraînant une accumulation de GMPc intracellulaire et une relaxation des fibres musculaires lisses. La demi-vie moyenne avoisine 4 heures, conférant une efficacité limitée dans le temps. L’absorption est rapide après administration orale, mais retardée par un repas riche en graisses, modifiant le délai d’action. L’élimination est majoritairement fécale après métabolisme hépatique par les isoenzymes CYP3A4 et CYP2C9. Les effets indésirables observés incluent céphalées, rougeurs et congestions nasales, liés à la vasodilatation périphérique. Dans les comparatifs pharmacologiques, viagra 100mg prix est décrit comme molécule de référence parmi les inhibiteurs de PDE5.
Barc16
IFA AGRICULTURAL CONFERENCE ON MANAGING PLANT NUTRITION 29 June - 2 July 1999, Barcelona, Spain THE IMPACT OF BIOTECHNOLOGY AND GENOMICS ON THE FERTILIZER INDUSTRY David A. Lightfoot
Southern Illinois University, United States
THE IMPACT OF BIOTECHNOLOGY AND GENOMICS ON THE FERTILIZER INDUSTRY1 David A. Lightfoot
Southern Illinois University, United States
1. Summary Problem to be solved: Agriculture impacts the environment. The Biotechnology and Genomics industries are seeking novel biotechnological methods to reduce negative impacts and increase positive impacts. Accelerated breeding using DNA markers and transgenes that improve crop plants are being used to promote positive modifications to current crop production systems. The Biotechnology and Genomics industries focus on those growers most likely to be profitable in the future free market agriculture. That group of growers control their inputs closely to match outputs and only lose about 10- 15% of input fertilizers compared to 50% losses measured from inefficient growers. Therefore, in Illinois the IEPA estimates that about 5% of growers cause 50% of nutrient pollution by: not using non-conservation tillage methods; overfertilizing relative to yield (1.3 lb of N per bushel vs. 1.1 lb per bushel as recommended); fall fertilization instead of spring fertilization; and simply being part of a watershed. Mandating appropriate cultural practices is a matter for public policy. Nutrient losses from non-point sources is a public health issue. The Biotechnology and Genomics industries can assist by reducing the 10-15% losses of the best growers through modifying crop plants to increasing yield per acre; improving yield stability (drought tolerance) and increasing the percentage of nutrients harvested (yield per input). At SIUC we have focused on generating GMOs with genes capable of improving plant nitrogen assimilation and metabolism; on improving non-GMO crop yield and yield stability through marker aided breeding; and latterly on application of genomics to both. 2. Introduction
A. Environmental Impacts of Crop production Yield, protein and nitrogen contents of plants are often limited by the uptake of nitrogen (N), phosphorous (P) and potassium (K) from the soil. Cereal crops grown for protein content and yield require large quantities of fertilizers (60-400 kg/ha) and are occasionally treated with nitrification inhibitors (NI). Yield increases may be large, but the fertilizers applied are not all assimilated by the crop and represents lost producer income. Nitrogen that is not assimilated into plant material will contribute to several undesirable environmental effects but especially nitrate and nitrite contamination of ground water (Benes et al., 1987). Diffuse nitrates in groundwater could seriously compromise drinking water quality for over 50 million Americans (Lee & Nielsen, 1987) and 25% of Illinois residents (see Kovacic et al., 1996; David et al., 1997). The removal of nitrates from drinking water presents economic and technical difficulties (Mellor et al., 1992). Diffuse nitrates carried to the sea by ground water can promote toxic algal blooms in oceanic waters, disrupting fisheries and tourism (Smayda, 1989; Cherfas, 1990; Burkholder et al., 1992). In addition the plants grown with nitrate nutrition can contain high tissue concentrations of nitrate and nitrite which are nutritionally undesirable (Marschner, 1995). Elevated levels of nitrates and nitrites in both foodstuffs and drinking water are implicated in several healthcare problems. Total nitrate intake determines the rate of formation of nitrosamine, a carcinogen implicated in the 1 Paper presented at the IFA Agricultural Conference on Managing Plant Nutrition, 29 June-2 July 1999, Barcelona, Spain
etiology of many human cancers (Tannenbaum et al., 1978; Mirvish, 1985, Moller et al., 1990). Dietary nitrates derived from vegetables and water can represent 70-85% of total intake (Mirvish, 1985; Moller et al., 1990). Infants fed with nitrate or nitrite-containing foods or water can develop methemoglobinemia (Marschner, 1995). Canned food high in nitrates can cause detinning of containers within a few months (Farrow, 1972). Forages high in nitrates can be toxic to ruminants and diffuse nitrates in drinking water may promote abortion (Prins, 1983). B. Genetic Manipulation of Crop Plants
Genetic improvement of crops has underlain yield that consistently increased by 2-4% per year in the major crops since 1930 (Duvik, 1990). Concomitant with yield the plants need for fertilization has increased. A consequence of yield improvement has been a restricted genetic base and increasing yield instability on a year by year basis (Duvick, 1993). Instability has resulted in localized failures to harvest the fertilizer inputs as yield and there is a need to avoid losses in these situations. Genetic improvement of elite germplasm will increasingly rely on transgenes and GMOs in future. Genes that can increase yield and yield stability are being sought (Lightfoot et al., 1998; Willmitzer et al., 1998). Understanding how these genes work will teach how to maximize the benefits of their use for the growers and public. Plant growth and composition can be altered by nitrogen nutrition (Magalhaes et al., 1990; Doehlert et al., 1991; Marschner, 1995). Manipulation of plant nitrogen assimilation by toxins (Knight and Langston-Unkefer, 1988; Osuji and Cuero, 1992), mutation (Melo-Olivera et al., 1996) or transgenes (Temple et al., 1993) can alter growth, composition and abiotic stress resistance (Ameziane et al., 1999; Schmidt et al., 1999). These effects reflect the interrelationship of nitrogen metabolisms with all other plant processes. We use isogenic transgenic plants to elucidate the mechanisms coordinating plant growth development and composition. 3. Examples of Genomics and Biotechnology Products Impacting the Fertilizer Industry I will discuss examples of GMO crops and normal genetic improvements achieved through breeding that can impact the fertilizer industry A. GMO Crops Crops that are altered in nutrient biochemistry are being produced including GDH (glutamate dehydrogenase) and phytate reduced crops. I. GDH The strategy for corn improvement (Lightfoot et al., 1988) has been to provide corn with an gene that produces an enzyme (NADP-dependent glutamate dehydrogenase) from soil bacteria (Figure 1) that is found in no plant species (Lightfoot et al., 1998). A similar approach is being followed with the Chlorella gene (Schmidt et al., 1999). The gene, gdhA, comes from an organism that grows well on ammonium but not nitrate nitrogen and is responsible for that organism's vigorous growth on ammonium nitrogen. In plant the enzyme causes increased N assimilation, increased yield (and yield stability), resistance to Liberty herbicide and resistance to ammonium "burn" of corn during fertilization (Figures 2-3; Long et al., 1996). The gene provides about 10% greater yield from the same fertilizer input. However, the gene performs best with high fertilizer inputs. Therefore, the gene can contribute to both public health and grower profitability in Illinois and the USA. It is an example of green biotechnology. For the growers and breeders we are currently identifying the most promising events among the current set of transformants. To advance scientific knowledge and maximize the benefits that the gene can provide we are undertaking a combined physiological and genomic analysis. II. Reduced Phytate A genotype is envisioned that will take up P and yield well with lesser P inputs, accumulate P in vegetative tissues contain less P in grain but in a form suitable for non-ruminant digestion. Most of these goals will be achieved by low phytate crops (Verwoerd et al., 1995) where the added benefit of increased feed value and
reduced hog waste disposal costs also accrue. Such technology may reduce both the chemical N and P requirements of growers. III. Future GMOs (yield, nodules)
Targets for GMO crops in future will be identified with the gene discovery mechanisms provided by genomics and biotechnology. Using genomics the genetics of complex traits can be studied. Therefore, projects that were previously intractable will be addressed. One example is the ability of plants to form functional nodules for symbiosis with Rhizobium species (Men et al., 1999). If genomics can be used to identify a 100-200 gene complement for this characteristic the technology exists to enable transfer from legume crops to cereal crops. Such technology might drastically reduce N input requirements worldwide. Another example of an intractable character is yield, a trait that is always multi-genic and inherent to elite crops. If genomics can be used to identify a 50-100 gene complement for this characteristic the technology exists to enable transfer from among cereal crops. As noted earlier increased yield requires increased fertilization. A third intractable trait is the question of what causes the large differences in fertilizer use efficiency that we observe among cultivars within crop species and between species. If genomics can be used to identify a 10-20 gene complement for this characteristic the technology exists to enable transfer among cultivars or between crop species. Selection could be applied toward efficiency or inefficiency. B. Marker Assisted Breeding of Crops A. Marker assisted selection for yield and disease resistance
Selection for increased yield potential is the ultimate goal of all plant breeding programs. Most of the yield increases over the past 60 years have been due to genetic advances by intercrossing existing varieties (Duvick et al., 1992). However, yield is a multigenic trait and therefore the yield potential of lines derived by inter-crossing is difficult to predict without extensive field testing. Selection for yield relies on repeated selection for components of the yield by the breeder over a decade. Often, selection begins at the F5 with selection for maturity group and growth habit. This will reduce 2000 lines to 200 within a typical population. Selection for the desired disease resistances in the greenhouse can reduce the lines to be analyzed to 50. After that, repeated field testing in multiple environments over 8-9 years will identify one or two lines with better yield performance than existing varieties. Overall yield advances of 2% per year have been achieved for new soybean varieties over the last 30 years by this approach (Duvick et al., 1992). However, yield increases for new corn varieties (and other outcrossing crops where genetic diversity is not limiting) have been double this at 4% per year over the same period. Two avenues to increasing the rate of increase of crop yields exist. Both require the deployment of marker assisted breeding methods. The first, using current adapted germplasm, is to increase the number of lines with high yield potential in the later stages of some breeding programs. This might be achieved with molecular markers by increasing the abilities of breeders to identify lines with high yield potential by early selection. The second, using exotic germplasm, is to discover new genes and alleles that can increase yields of new adapted varieties as well as increase their genetic diversity. This might be accelerated by marker assisted discovery of the new genes and alleles in exotic germplasm (Tanksley et al., 1996). At SIUC we are engaged with both approaches. By identifying molecular markers associated with high yields in genetically diverse lines and under different cropping environments we can give plant breeders powerful new tools for identifying new higher yielding varieties (Figure 4). Typically using two markers the breeder becomes four fold more effective in advancing yield. Therefore both yield and fertilizer input requirements should increase well above the current 2-4%. C. Genomic Techniques
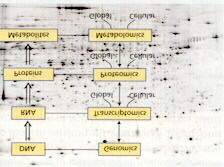
Genomic techniques will accelerate all approaches to crop improvement. Genomics is the science derived by parallel processing biological data, that is, the application of computing (bioinformatics, automation) to biology (Hieter and Boguski, 1996). In genomics we study everything at once as opposed to the one gene, one protein, one effect paradigm of the earlier approaches. Taxonomy of genomics derives from the element
studied (Figure 5), genome-genomics, proteins proteomics, metabolites to metabolomics. Genomics is split into structural (chromosomal) and functional (transcriptomics) I. Applications of Structural Genomics
Chromosomes can be deconstructed and cloned in large insert libraries then fingerprinted to allow reconstruction of clone order to generate the originating genome (Figure 6; Meksem et al., 1998; 1999). Uses for arrayed libraries are many but the isolation of genes underlying QTL for use in GMOs and the generation of ideal markers for marker aided breeding are the two most immediately applied to fertilizer response and efficiency. Sequencing of the whole genome will also be accelerated by this technology. II. Applications of Functional Genomics
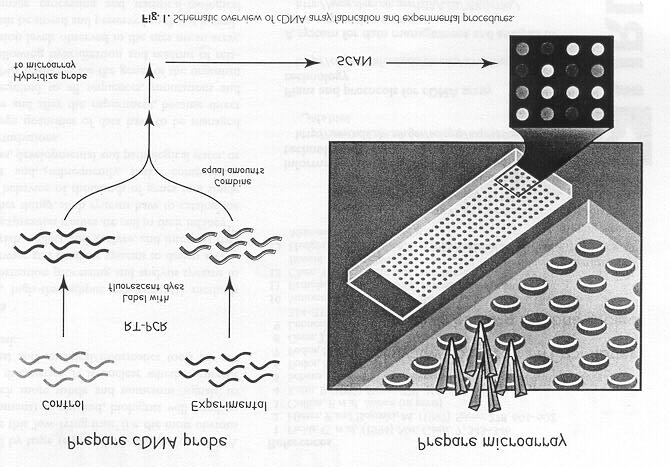
Sequencing of the every gene as a transcript (EST) is now commonplace from which the sequence of every protein can be inferred (DeRisi et al., 1997). Using automation and miniturization every gene from a crop can be arrayed on a microscope slide and the response to a treatment or transgene studied (Figure 7). The methods are being applied to GDH crops and crop germplasms that contrast for responses to nutrients. Using these techniques new genes involved in well established biochemical pathways have been discovered. These genes are targets for manipulation in GMOs or by breeding. III. Applications of Proteomics Proteomics analyses every protein in a cell simultaneously (Figure 8). It is important because there is often poor correlation between gene array data and protein abundance suggesting important genes will be missed if studying only the transcript abundance with arrays. The technology includes 2-D Gels and advanced methods of protein identification (MALDI-TOF) based on DNA sequence knowledge. IV. Applications of Metabolomics
The ultimate effect of a transgene or breeding line modification will ultimately be expressed as a change in metabolite concentration. Metabolomics seeks to measure such low molecular weight compounds rapidly in parallel. Integration - Surfing the “Web”
Combining approaches will allow identification of important new genes for fertilizer response and efficiency within the web that determines crop responses. The methods will be used to maximize the benefits of existing transgenes and prove equivalence between GMOs and non-GMOs. Conclusions
The impacts can be both negative and positive in effect on fertilizer sales worldwide. However, even apparent negative technology may positively impact the fertilizer industry because any technology that facilitates the economic development of biofuel crops could double crop production requirements for every 10% of fossil fuel replaced. Therefore, technologies that reduce fertilizer inputs per unit yield might increase the total market dramatically. References
Ameziane R. Bates, R. Long L.M., Vidal, M.E., Lightfoot, D.A. 1999. Expression of the Escherichia coli glutamate dehydrogenase gene in Nicotiana tabacum causes resistance to the herbicide phosphinothricin and exogenous ammonium chloride. Nature Biotechnology (accepted). Benes, V., Pekny, V., Skorepa, J. & Vrba, J. (1989). Impact of diffuse nitrate pollution sources on groundwater quality - some examples from Czechoslovakia. Environ. Health Perspect 83: 5-24.
Burkholder, J.M., Noga, E.J., Hobbs, C.H. & Glasgow Jr., H.B. (1992). New 'phantom' dinoflagellate is the causative agent of major estuarine fish kills. Nature 358:407-410. Cherfas, J. (1990). The fringe of the ocean - under siege from land. Science 248, 163-165. David, M.B., Gentry, L.E. Kovacic,D.A., Smith K.M. (1997). Nitrogen balance in and export from an agricultural watershed. J. Env. Qual 26:1038-1048. DeRisi J.L., Vishwanath R., Brown P.O. Exploring the metabolic and genetic control of gene expression on a genomic scale. Science 278:680-687. (1997). Doehlert, D. and Lambert, R.J. (1991). Metabolic Characteristics associated with starch, protein and oil deposition in developing maize kernels. Crop Sci. 31:151-157. Duvick, D.N. 1991. Progress in Conventional Plant Breeding. Springer Verlag, Amsterdam. Farrow, R.P., Johnson, J.H., Gould, W.A. & Charbonneau, J.E. (1971). Detinning in canned tomatoes caused by accumulations of nitrates in the fruits. Food Sci. 36:341-345. Keeney, D.R. and Nelson, D.W. (1982). Nitrogen Inorganic form. In A.L. Page et al. (ed) Methods of soil analysis. Part 2 Agronomy 9: 643-698. Hieter P and Boguski M. Functional genomics: Its all in how you read it. Science 278:601-603. Knight, T.J. & Langston-Unkefer, P.J. (1988). Enhancement of symbiotic dinitrogen fixation by a toxin-releasing plant pathogen. Science 241:951-953. Kovacic,D.A., David, M.B., Gentry, L.E. (1996). Grassed detention buffer strips for reducing agricultural nonpoint-source pollution from tile drainage systems. Proceedings of the sixth annual conference of the Illinois groundwater consortium, March 27-28, 1996. Carbondale: IGC, Southern Illinois University at Carbondale. pp88-97. Lee, L.K. & Nielsen, E.G. (1987). The extent and costs of groundwater contamination by agriculture. J. Soil Water Conserv. 42:243-248. Long L.M., Vidal, M.E., Lightfoot, D.A. 1996. Plants containing the bacterial gdhA gene and methods of use thereof. US Patent pending issue. Lightfoot, D.A., Baron, A.J. & Wootton, J.C. 1988. Expression of Escherichia coli glutamate dehydrogenase in the cyanobacterium Synechococcus PCC6301 causes ammonia tolerance. Plant Molec. Biol. 11:335-344. Lightfoot, D.A. Bernhardt, K., Ameziane R. Bates, R. Long L.M., 1998. Expression of the Escherichia coli glutamate dehydrogenase gene in Nicotiana tabacum and Zea mays increases yield. Proceeding of the Illinois Corn Breeders School 34:118-149 Magalhaes, J.R. Ju, G.C. Rich P.J. and Rhodes, D. (1990). Kinetics of 15NH +
Physiol. 94:647-656. Meksem K., Zhang H-B., and D.A. Lightfoot. 1998. A plant transformation ready bacterial artificial chromosome library for soybean: Applications in chromosome walking and genome wide physical mapping. Soybean Genetics Newsletter 25:139-141. Meksem K., Zhang H-B., and D.A. Lightfoot. 1999. Two plant transformation ready bacterial artificial chromosome libraries for soybean: Applications in positional cloning of QTL. Theoretical and Applied Genetics. In press.
Mellor R.B., Ronnenberg, J. Campbell, W.H. & Diekman, S. (1992). Reduction of nitrate and nitrite in water by immobilized enzymes. Nature 355:717-719. Men A., Vasile C-A., Andersson I., Bhattacharrya M., Meksem K., Lightfoot, D.A., Gresshoff P.M. 1999. Genomic analysis of the soybean supernodulation mutation nts-1. Genome In press. Marchner, H. (1995). Mineral Nutrition of Higher Plants. Academic Press, London, UK. Mirvish S.S. (1985) Gastric cancer and salivary nitrate and nitrite. Nature 315:461-462 Moller, H., Landt, J., Pederson, E., Jensen, P., Autrup, H., & Jensen, O.M. (1990). Endogenous nitrosation in relation to nitrate exposure from drinking water and diet in a Danish rural population. Cancer Res. 49:3117-3121. Osuji, G.O. & Cuero, R.G. (1991). Regulation of ammonium ion salvage and enhancement of storage protein contents of corn, sweet potato and yam tuber by N-(carboxymethyl)chitosan application. J. Agric. Food Chem. 40:724-734. Prins, W.H. (1983). Effect of a wide range of nitrogen applications on the herbage nitrate content in long term fertilizer trials in all-grass swards. Fert. Res. 4:101-113. Sakakibara, H., Fujii, K. and Sugiyama, T., (1995). Isolation and characterization of a cDNA that encodes maize glutamate dehydrogenase. Plant Cell Physiol. 36:789-797. Smayda, T.J. (1989. In Novel Phytoplankton Blooms ed Cosper, E.M. et al. Springer, New York, pp449-484. Schmidt RR., et al., 1999. Polypeptides and polynucleotides relating to alpha and beta subunits of glutamate dehydrogenase and methods of use. US patent # 5879941. Tanksley S.D. and Nelson J.C. 1996. Advanced Backcross QTL analysis: A method for the simultaneous discovery and transfer of valuable QTL from unadapted germplasm to elite breeding lines. Theor. Appl. Genet. 92:191-203. Tannenbaum, S.R., Fett,D., Young, V.R., Land, P.D. & Bruce, W.R. (1978). Nitrite and nitrate are formed by endogenous synthesis in the human intestine. Science 200:1487-1491. Temple, S.J., Knight, T.J., Unkefer, P.J. and Sengupta-Gopalan, C. (1993). Modulation of glutamine synthetase gene expression in tobacco by the introduction of an alfalfa glutamine synthetase gene in sense and antisense orientation: molecular and biochemical analysis. Mol. Gen. Genet 236:315-325. Verwoerd, TC. van Paridon, PA., van Ooyen J.J. van Lent JWM., Hoekma A. and Pen J. 1995. Stable accumulation of Aspergillus niger phytase in transgenic tobacco leaves. Plant Physiology 109:1199-1205.
Figure 1: N Assimilation Differs In Plants and Microorganisms Height 1 2 0 gm/plant 40 H e i g h t K e r n e l S t o v e r Selections 1996 1997 1998
NUVARING NAME OF THE MEDICINAL PRODUCT NuvaRing® 2. QUALITATIVE QUANTITATIVE COMPOSITION NuvaRing contains 11.7 mg etonogestrel and 2.7 mg ethinylestradiol. The ring releases etonogestrel and ethinylestradiol at an average amount of 0.120 mg and 0.015 mg, respectively per 24 hours, over a period of 3 weeks. For excipients, see 6.1. 3. PHARMACEUTICAL NuvaRing is flexible,
Prolonged Methylprednisolone Treatment Suppresses Systemic Inflammation in Patients with Unresolving Acute Respiratory Distress Syndrome Evidence for Inadequate Endogenous Glucocorticoid Secretion and Inflammation-induced Immune Cell Resistance to Glucocorticoids G. Umberto Meduri, Elizabeth A. Tolley, George P. Chrousos, and Frankie Stentz Memphis Lung Resea
 Figure 1: N Assimilation Differs
Figure 1: N Assimilation Differs Selections
Selections