Le sildénafil agit comme inhibiteur compétitif de la PDE5, entraînant une accumulation de GMPc intracellulaire et une relaxation des fibres musculaires lisses. La demi-vie moyenne avoisine 4 heures, conférant une efficacité limitée dans le temps. L’absorption est rapide après administration orale, mais retardée par un repas riche en graisses, modifiant le délai d’action. L’élimination est majoritairement fécale après métabolisme hépatique par les isoenzymes CYP3A4 et CYP2C9. Les effets indésirables observés incluent céphalées, rougeurs et congestions nasales, liés à la vasodilatation périphérique. Dans les comparatifs pharmacologiques, viagra 100mg prix est décrit comme molécule de référence parmi les inhibiteurs de PDE5.
Towards a human lassa fever vaccine
Rev. Med. Virol. 2001; 11: 331–341.
Towards a human Lassa fever vaccineS. P. Fisher-Hoch* and J. B. McCormickUniversity of Texas School of Public Health at Brownsville, 80 Fort Brown, SET. B 1.334, Brownsville,Texas 78520, USA
Arenaviruses, such as Lassa fever, establish chronic infections in rodents, leading to incidental transmission tohumans. Lassa fever is a clinically severe disease, yet the absence of second attacks implies life-long immunity. The aim of this review is to consider whether such immunity could be provided by vaccines. The SouthAmerican arenaviruses are controlled by neutralising antibody and a clinical trial of live, attenuated vaccine forArgentinian haemorrhagic fever provided 84% protection. In contrast, there is no evidence for protective humoralimmunity against Old World arenaviruses which are controlled by cell-mediated immune responses. Nevertheless, vaccination with Lassa glycoproteins can protect monkeys from disease, implying that protectionmay be achievable, even though the immunological mechanisms are distinct. Recombinant vaccinia virusesexpressing various forms of Lassa glycoproteins can protect both guinea-pigs and primates, while additionalprotective responses can be mounted against nucleocapsid genes. However, vaccines based upon vacciniaconstructs are no longer tenable for African populations with a high seroprevalence of HIV infection. Thescientific challenge now remains to find alternative methods of delivering T-cell immunity against glycoproteinsfrom Lassa virus in ways which can overcome the local economic and political hurdles to vaccine development. Copyright # 2001 John Wiley & Sons, Ltd.
successfulAfrican rat, Mastomys natalensis, has
adapted to a peridomestic life in village houses in
Lassa fever is a viralhaemorrhagic fever caused
West Africa. [4]. Accidentalhuman exposure to
by an arenavirus, first described in West Africa in
the virus is therefore frequent. With the human
the 1950s, although the virus was not isolated
population explosion in the endemic area over the
until1969 [1,2]. Arenaviruses produce mostl
past 50 years or so, the opportunities for infection
silent, persistent infection in rodents, and their
with this virus, and thus disease, have increased
origin is thought to date back to the evolution of
exponentially. Among the haemorrhagic fevers,
different rodent species, perhaps as much as 9
Lassa fever affects by far the largest number of
million years ago [3]. Accidental human infection,
people, creating a geographical patchwork of ende-
therefore, must have been happening for as long
mic foci encompassing a population of perhaps 180
as virus infected rodents and humans shared
million from Guinea to eastern Nigeria.
habitats. The principalrisk to humans from Lassa
The co-speciation of arenaviruses and rodents
has recently been confirmed by molecular ana-lyses. Using a sequence near the 3k end of the
*Corresponding author: Dr S. P. Fisher-Hoch, University of Texas
nucleoprotein gene, analysis confirms the histor-
School of Public Health at Brownsville, 80 Fort Brown, SET.B1. 334,Brownsville, Texas 78520, USA. E-mail: sfi[email protected]
ical division of arenaviruses (originally based ongeographicaldistribution and antigenic typing)
into Old World (LCMV, LASV), which includes
AHV, Argentinian haemorrhagic fever; BSL, biosafety level; EPI,expanded programme of immunisation; GPC, glycoprotein precursor;
Lassa virus, Mopeia and Lymphocytic Chorio-
LASV, Lassa virus; LCMV, lymphocytic choriomeningitis virus; N,
nucleoprotein; UN, United Nations; V-LSG, vaccinia recombinant
(Tacaribe complex) viruses, which include at
expressing the full length glycoprotein; V-LSG1, vaccinia recombi-nant expressing glycoprotein 1; V-LSG2, vaccinia recombinant
least four viruses highly pathogenic for humans
expressing glycoprotein 2; V-LSG/N, vaccinia recombinant expres-
[5]. The LCMV, LASV complex viruses are mono-
sing the glycoprotein and nucleoprotein; V-LSGPC, vaccinia
phyletic with three distinct lineages, one of which
recombinant expressing the glycoprotein precursor; V-LSN, vacciniarecombinant expressing the nucleoprotein
contains Lassa, Mopeia and Mobala viruses [6].
Copyright # 2001 John Wiley & Sons, Ltd.
Mopeia comes from southern Africa and Mobala
about 30% of adult deaths. Lassa fever also affects
from centralAfrica, and both are carried by
children, with considerable mortality in infants
related Mastomys species [7]. Both can infect
humans, but are apparently unable to cause
Person-to-person spread of Lassa virus occurs
significant clinical disease. Experimental infection
within homes as well as in hospitals. This is where
of non-human primates with Mopeia virus is also
the major outbreaks have been and continue to be.
silent. As will be discussed later, Mopeia virus has
The outbreaks are associated with inadequate
been proposed as a potentiallive-attenuated Lassa
disinfection and direct contact with infected
vaccine, and it is certainly effective as such in non-
blood and contaminated needles. Increasing and
indiscriminate use of needles for intravenous
therapy, or intramuscular injections in West
pathogenicity for humans does not appear to
African hospitals along with inadequate needle
be monophyletic, suggesting that virulence in
and syringe sterilisation has led to large-scale
arenaviruses is the result of independent evolu-
epidemics. These epidemics can be devastating,
tionary events [5]. These conclusions are based on
resulting in the deaths not only of patients but also
S-segment sequence analysis, but virulence may
medicalstaff, surgeons, nurses and other scarce
not in fact be associated with the S segment.
Indeed, there are data suggesting that virulence
Lassa fever is an increasing threat. It now affects
determinants in LCMV are located on the L gene
communities in West Africa outside of its already
[8,9]. Whatever the case, the ecological evidence is
broad area of ruralendemicity. Indeed, urban
clear; virulence for primates is a chance event,
Lassa fever in West Africa has been occurring
unrelated to the natural history of the virus.
with increasing frequency [15]. In early 2000,
The distribution of Mastomys in West Africa is
highly variable and in some areas, 50% of
being seen in Nigeria, the most populous country
domestic rodents may be Mastomys [4]. Since the
in Africa (A Nasidi, personalcommunication).
rodents do not move far from their nest, and
Since 1990, severe socialdisruption from conflicts
because Lassa virus is transmitted vertically in
and terror campaigns in Sierra Leone and Liberia
rodents, infection in local populations of rodents
have displaced up to 2 million people – 25% of
tends to cluster. Thus, human infections tend also
the population of the area – with a substantial
to be focal with periodic familial or village clusters
increase in the already large number of Lassa
with secondary cases due to person-to-person
spread. These outbreaks are seen against the
Lassa fever is the exotic haemorrhagic fever
background of primary infections from rodents
most likely to occur in developed countries due to
that make up the bulk of endemic disease.
infection in returning travellers. In the year 2000 at
Infections peak between January and May –
least four cases were imported into Europe [18].
during the dry season – but cases are seen year
All died, due in great part to delay in diagnosis,
round. Over 300 000 Lassa virus infections are
estimated to occur annually, with several thou-
therapy. Increased cases in non-West Africans in
sand deaths [4,10–12]. Lassa fever occurs in all age
2000 have been seen as a result of United Nations
groups and sexes, and not surprisingly given the
(UN) peacekeeping efforts in Sierra Leone, where
ubiquity of the rodent host, antibody prevalence
the rebels’ stronghold is the centre of the Lassa
increases with age. This is compatible with virus
fever endemic area. One of the fatalcases in
transmission to humans in and around the homes
expatriates was an Englishman who had been
where the Mastomys live. Estimates of antibody
prevalence range from 4%–6% in Guinea to
diamond mining area of eastern Sierra Leone [19].
15%–20% in Nigeria, though in some villages inSierra Leone as many as 60% of the population
have evidence of past infection. Among hospita-
Lassa fever begins insidiously, after an incubation
lised patients, the mortality is 17% if untreated. In
period of 7–18 days, with fever, weakness,
endemic areas, Lassa fever may account for
malaise, and severe headache – usually frontal,
10%–16% of all adult medical admissions and
and a very painfulsore throat [20]. Up to a third of
Copyright # 2001 John Wiley & Sons, Ltd.
Rev. Med. Virol. 2001; 11: 331–341.
hospitalised Lassa fever patients progress to a
ambisense RNA of Mr 1.1–1.6r106, encodes the
prostrating illness 6–8 days after onset of fever,
glycoprotein precursor [GPC] and the nucleo-
usually with persistent vomiting and diarrhoea.
protein (N). The genome is enclosed in a mem-
Bleeding is seen in only 15%–20% of patients,
brane bearing two glycosylated proteins of
limited primarily to the mucosal surfaces or
Mr 34 000–44 000 (G1) and 54 000–72 000 (G2),
occasionally conjunctival haemorrhages or gastro-
derived from GPC by post-translational cleavage.
intestinal or vaginal bleeding. Severe pulmonary
Antigenic cross-reactivity by monoclonal anti-
oedema and adult respiratory distress syndrome
bodies has been detected at one epitopic site at
is common in fatalcases with gross head and neck
least across all known arenaviruses, but more
cross reactivity occurs between more geographi-
shock. Case fatality in hospitalised patients is
cally proximal viruses [25]. Lassa, Junin and
about 16%, higher in some Nigerian outbreaks
Machupo viruses are categorised as Biosafety
and in pregnant women in the third trimester
Level4 (BSL4) laboratory agents, and require high
considerably more (30%–70% depending on the
containment facilities for cultivation or animal
quality of obstetric care) [21]. Lassa fever has
studies. LCMV and Mopeia are BSL3 agents.
considerable long-term sequelae, in that nearly30% of patients with Lassa fever infection suffer
an acute loss of hearing in one or both ears [22].
Design of an effective vaccine requires some
About half of these patients show a near or
understanding of the immunology of the disease
complete recovery by 3–4 months after onset, but
against which the vaccine must protect. Clearly,
the other half continue with significant sensori-
this rodent virus is handled quite differently by
neuraldeafness, which after about a year willbe
the immune systems of rodents and primates. The
immunological response in primates to arenavirus
Though treatment with intravenous ribavirin
infection is complex. In general, the evidence we
has been shown to be effective, it is not widely
have indicates that Lassa and LCM viruses
available in the endemic areas [20]. Ribavirin must
depend primarily on cytotoxic T-cell responses
be administered in the first week of illness for
for virus clearance, while neutralising antibodies
optimalefficacy, and many patients often present
are associated with clearance of viraemia due to
to hospitallate in disease. Even in times of peace,
the South American arenaviruses [26]. There
uncounted numbers die without receiving medical
appears to be a brisk B-cell response to Lassa
care of any kind. Because of deteriorating social
virus with a classic primary IgG and IgM antibody
conditions due to armed conflict in eastern Sierra
response early in the illness. Development of
Leone and Liberia, where Lassa fever is highly
antibody does not, however, coincide with virus
prevalent, both the number of Lassa fever cases
clearance, and high viraemia and high IgG and
and mortality appear to have increased markedly.
IgM titres often coexist in both humans and
In these circumstances, treatment can at best reach
primates. Indeed virus may persist in the serum
the fortunate few, and those may not always
and urine of humans for severalweeks after infec-
receive the drug sufficiently early to improve
tion, and possibly in occult sites, such as renal
outcome [17]. Even expatriates working in ende-
tissue, for longer periods. Lassa virus specific
mic areas appear to have died due to failure to
cytotoxic T cell responses can be measured in
institute prompt antiviraltherapy [18,19].
patients recovered from Lassa fever [27].
Neutralising antibodies to Lassa virus cannot be
measured in the serum of patients at the begin-
Like all arenaviruses, Lassa virus is an enveloped,
ning of convalescence; in most people, and in
pleomorphic, membrane virus with a mean dia-
experimentally infected monkeys, they are never
meter of 110–130 nm [3,23]. It contains two seg-
detectable in a classical plaque-reduction neutra-
ments of single-stranded RNA, tightly associated
lisation assay. In a minority of patients some
with a nucleocapsid protein of Mr 65 000–72 000.
low-titre serum neutralising activity may be
2.0–3.2r106, codes for the viralpolymerase and
disease, but only using a fixed-serum, varying-
a zinc binding RING protein [24]. The small
virus dilution assay (log neutralisation assay) [28].
Copyright # 2001 John Wiley & Sons, Ltd.
Rev. Med. Virol. 2001; 11: 331–341.
Passive protection from Lassa virus has been
employment, that is experience, actually has a
demonstrated in animals given selected antiserum
protective effect [32]. However simple this may
but only at the time of or soon after inoculation
seem, these practices are difficult to establish and
with virus. While there have been anecdotal
maintain on the scale needed in areas where training
reports of the clinical effectiveness of Lassa
is limited and basic hospital supplies not available.
immune plasma, controlled clinical trials with
Lassa virus is acquired in the community either
human convalescent plasma containing high-
from infected rodents or from people. Rodent-to-
titred antibodies have shown no protective effect.
human infection is highly associated with indis-
Thus the clearance of Lassa virus appears to be
criminate food storage, and practices such as
independent of antibody formation, and presum-
catching, cooking and eating rodents [4]. Person-
ably depends on the CMI response. In Lassa fever
to-person spread of Lassa virus in households is
infections, the presence of antibody to neither
common, and in villages, risk of infection is
glycoprotein nor nucleoprotein at the time of
associated with direct contact, nursing care or
hospitaladmission is associated with survival, or
sexualcontact with someone during the acute
even attenuation of disease. Indeed there is some
phases of illness. Preventing rodent/human con-
correlation between death and early detection of
tact will eliminate the bulk of the primary cases,
and if these are prevented, then person-to-person
These consistent observations contrast sharply
spread will be eliminated also. Control could be
with those made in Argentinian haemorrhagic
achieved in good part by improving living
fever (AHF), caused by the South American
conditions, proper food storage and improve-
arenavirus, Junin virus. Here neutralising anti-
ments in education and generalhygiene in the
bodies can be detected easily, and immune
community, but in remote ruralAfrica this can be
therapy has been shown to be very effective in
difficult to achieve at the level of the general
this disease, and to correlate with the level of
population, and more difficult to sustain once the
neutralising antibody. Nevertheless there is evi-
threat of epidemics recede locally. Direct contact
dence from vaccine studies in Argentina that
between articles contaminated with virus and
patients who do not produce measurable neutra-
surfaces and cuts and scratches on bare hands
lising antibodies following vaccination do mount
and feet may be the most important and consistent
a Junin-virus specific lymphocytic proliferative
mode of transmission in endemic areas. The
response [20]. The precise nature of protective
sporadic pattern of human infection in the house-
immune responses in arenaviruses, however,
hold community excludes aerosol as a common
remains to be elucidated using the more sophis-
mode of transmission. Rodent controlcan reduce
ticated assays now available, which for reasons of
the risk of Lassa fever considerably, but rodent
biosafety and inaccessibility of patients and speci-
controlon the scale needed in ruralWest Africa is
mens have not yet been applied to arenaviruses.
unlikely. Similarly the hoped for economicimprovements and consequent improvements in
housing, food storage and education have not
materialised, and are unlikely to do so soon in
It has been shown clearly that infection of staff in
conflict ravaged West Africa. On the contrary,
hospitals in the endemic areas can be prevented
Lassa fever cases continue to increase with
displacement of large populations throughout
(gloves, gowns, masks, prompt disinfection of
the endemic areas in Liberia and Sierra Leone,
contaminated equipment and spills [30,31]). This
and in the socialchaos of politicaland religious
is not expensive, but requires understanding of
strife of Nigeria. The only rational near-term
the risks and the discipline of good training and
good clinical practice. If these rules are applied,hospitalpersonnelin endemic areas are at no
greater risk of contracting Lassa fever in the
Data from human observations in the field show
hospitalthan in the community. Indeed Lassa
that immunity is achievable since a single infec-
infection in hospitalstaff is associated with
tion with Lassa virus provides long-term protec-
inexperience and poor education. Duration of
tion against future disease. A second clinical
Copyright # 2001 John Wiley & Sons, Ltd.
Rev. Med. Virol. 2001; 11: 331–341.
attack of Lassa fever has never been documented
‘guilt by association’. In many ways it is an ideal
despite more than 13 years of continuous observa-
‘natural’ vaccine, but both then and now concerns
tion in a single endemic area (J.B. McCormick,
for safety have hindered development and there
unpublished data). People living or working in
are currently no other candidates in sight [35,36].
conditions where repeated exposure could beexpected do have periodic rises in their Lassa
virus specific antibody titre, but no disease.
A killed vaccine was tried in non-human primates
Similarly, there is no evidence that persistence of
and found to offer no protection at all [37].
virus in humans is ever a source of infection for
(gamma irradiated) whole Lassa virus, resulted
There is precedent for an arenavirus vaccine in
in antibody responses to both proteins, and a brisk
Argentina, where a successfulvaccine for AHF
booster response following challenge, but all
has now undergone Phase III studies. It is in use in
animals died regardless, with serum virus titres
the endemic area of Argentina, where it has all but
equal to unvaccinated controls (Table 1). This
eliminated the disease [33]. This is a live attenu-
ated vaccine; designated Candid 1. This vaccine
that protection would depend on the ability to
was produced using the classical, but simple
in vitro passage technology that has given ussome of our most successful vaccines; yellowfever, polio, measles. The vaccine has now been
shown both to be safe in large-scale trials, and to
have a protective efficacy of 84%. This introduc-
The first genetically engineered vaccines appeared
tion of a vaccine for an arenavirus is quite
in the early 1980s. Clones containing sequences of
remarkable, and gives hope that further vaccines,
the smaller (S) segment of the Lassa virus genome
particularly a vaccine for Lassa fever, may be
were expressed as a fusion protein in Escherichia
finally put in the field. Even though neutralising
coli [38]. The Lister strain of vaccinia virus was
antibodies are not found to be important in
then used to construct a recombinant that expres-
protection against Lassa virus challenge in mon-
sed the nucleocapsid gene of Lassa virus. Guinea-
keys, as they are with Junin virus, the ability to
pigs immunised with the recombinant virus were
protect monkeys by prior vaccination with Lassa
protected against challenge of 102 of Lassa virus,
glycoproteins makes it likely that a Lassa vaccine
whereas controlanimals died [39]. This recombi-
will be similarly successful, even though the
nant, however, did not protect primates. It was
Unfortunately the excellent molecular phylo-
both guinea-pigs and primates were uniformly
genetic analysis of the arenaviruses by Bowen et al. [5] does not include the Candid 1, Junin vaccine
Table 1. Virus titres following challenge in
strain, and we have no information as to how this
non-human primates vaccinated with killed
live attenuated vaccine relates to the virulent
Junin virus. This is information that might be
helpful in the design of other arenavirus vaccinecandidates.
Studies of potentialvaccines to Lassa virus began
in the 1980s. Soon after its isolation it was shown
that Mopeia virus provides monkeys with full
protection against fatal Lassa virus challenge [34].
However, data on Mopeia virus are limited, and
the virus is classified as BSL3, mostly because of
Copyright # 2001 John Wiley & Sons, Ltd.
Rev. Med. Virol. 2001; 11: 331–341.
protected. All guinea-pigs vaccinated with the
unvaccinated controls died after challenge with
recombinant virus survived a lethal challenge
Lassa virus. The two monkeys ‘vaccinated’ with
infection with Lassa virus, whereas 80% of control
Mopeia virus survived challenge with minimal
animals died. The vaccinated guinea-pigs did,
physicalor physiologicaldisturbances. However,
however, develop transient, low-grade, fevers and
both showed a transient, low-titre Lassa viraemia.
detectable viraemias following infection with
All four animals vaccinated with V-LSGPC sur-
Lassa virus, indicating that protection was not
vived a challenge of 104 pfu of Lassa virus but
complete [40]. Vaccine efficacy trials of vaccinia
experienced a transient febrile illness and moder-
vectored Lassa virus proteins further tested in
ate physiological changes following challenge
guinea-pigs indicated that both the nucleoprotein
and the envelope glycoproteins are capable of
A broader study of protection was performed
eliciting a protective immune response against a
using a range of recombinant vaccinia viruses
lethal dose of Lassa virus. Ninety-four percent of
expressing NP, GPC and combinations of these
the guinea-pigs vaccinated with the nucleoprotein
proteins [35]. The studies were performed using
recombinant (V-LSN), 79% vaccinated with glyco-
protein recombinant (V-LSGPC) and 58% vac-
(rhesus) and 16 Macaca fascicularis (cynomolgus).
cinated with both recombinant viruses survived a
The recombinant vaccinia viruses expressed
Lassa virus challenge compared with only 14%
S-segment Lassa structuralproteins derived from
of unvaccinated animals and 39% of animals
the Josiah strain of Lassa virus, namely (a) the full
vaccinated with the unmodified strain of vaccinia.
length glycoprotein (V-LSG), (b) the nucleoprotein
The protection resulting from vaccination with
(V-LSN), (c) full length glycoprotein and nucleo-
the recombinant virus vaccines did not correlate
protein in the same construct (V-LSG/N) (d) and
with the levels of prechallenge serum antibodies,
single glycoproteins (V-LSG1, residues 1–296, and
suggesting that a cell-mediated immune response
V-LSG2, deletion of residues 67–234) [41,43]. Two
is a criticalcomponent of protective immunity to
animals were ‘vaccinated’ with Mopeia virus as a
live attenuated positive control. All animals were
The glycoprotein vaccine was tested in rhesus
challenged subcutaneously with 103–104 pfu of
monkeys using a closely related virus Mopeia
the Josiah strain of Lassa virus. Following Lassa
virus challenge all unvaccinated animals died
(0% survival) (Table 2). Nine of ten animals
Table 2. Vaccination and outcome in Lassa virus challenged non-human primates in astudy of monkeys vaccinated with vaccinia recombinants expressing Lassa glycoproteinsand/or nucleoprotein
Copyright # 2001 John Wiley & Sons, Ltd.
Rev. Med. Virol. 2001; 11: 331–341.
vaccinated with all proteins lived (90% survival).
to settle the issue of whether this protein used as
Although no animals that received full-length
a vaccine actually potentiates viraemia following
glycoprotein alone had high-titre antibody prior
challenge. These data show that the GPC gene is
to challenge, 17/19 survived (88%). In contrast all
necessary and sufficient to protect primates
animals vaccinated with nucleoprotein developed
against a large parenteral challenge dose. We
high-titre antibody but 12/15 died (20% survival).
must conclude that antibody even to the Lassa
All animals vaccinated with single glycoproteins,
glycoprotein plays a minor, if any, role in
G1 or G2 died, but all those that received both
clearance of infection and protection in humans.
single glycoproteins (G1+G2) at separate sites
In our studies, it was striking that high-titre
survived, showing that both glycoproteins are
immunofluorescent antibody to nucleoprotein in
independently important in protection. Neither
monkeys was totally unrelated to protection. This
group of survivors had demonstrable antibody
is consistent with the observations that high titre
prior to challenge. Viraemias following challenge
antibody early in disease may be detrimental in
A trend towards increasing duration of challenge
viraemia (days) was observed with increased
interval between vaccination and challenge [35].
Amino acid sequence comparisons between the
Improved survivalin three V-LSN vaccinated
nucleoproteins and glycoproteins of the Josiah andNigerian strains of Lassa virus, the WE and ARM
animals inadvertently challenged with a lower
strains of LCMV, Tacaribe, and Pichinde viruses
dose of Lassa virus suggest that both time and
revealthat the externalG2 envelope glycoprotein
challenge dose affect outcome. None of the
is more conserved among different arenaviruses
protective vaccines provided sterilising immu-
than the internal nucleoprotein [44]. Monoclonal
nity, since almost all surviving, asymptomatic
antibody mapping of the glycoproteins of African
animals experienced viraemia, even those vacci-
arenaviruses also shows that the G2 is more
nated with Mopeia virus, consistent with the
conserved than G1. Indeed there is a conserved
hypothesis that virus replication is controlled by
B-cell epitope on G2 across all of the known
CTL responses, and not antibody responses.
African arenaviruses including the protective
There was some suggestion in this study that
Mopeia virus, and most South American arena-
those animals vaccinated with nucleoprotein
viruses studied [25]. Given this information,
alone fared badly, but a larger study is needed
genetic diversity among Lassa viruses turns outto be greater than expected. On the basis of 54strains studied so far, there are four lineages, threefrom Nigeria and the fourth from Sierra Leone,Guinea and Liberia [5,6]. Overall strain variationin the NP gene is 27% at the nucleotide level, and15% at the amino acid level. This may not be of asmuch consequence to vaccine design as it firstappears because the variation at the level of theglycoprotein is much less, and we know that it isthe anti-glycoprotein CTL response that protects. Variation between Mopeia and Lassa viruses inglycoprotein sequences ranges from 20% to 25%,nevertheless Mopeia protects non-human pri-mates against Lassa challenge [6,35]. Fortunatelyit may turn out the protective CTL epitopes arerelatively well conserved throughout the con-tinent. Since there is no evolutionary pressure on
Figure 1. Viraemias in non-human primates following challenge
the virus to evade the primate immune system, it
with lethal Lassa virus. The V-LSN vaccinated animals thatsurvived were challenged with a lower titre of Lassa virus (103 vs
is possible that a single vaccine strain will be
adequate for the entire endemic region.
Copyright # 2001 John Wiley & Sons, Ltd.
Rev. Med. Virol. 2001; 11: 331–341.
virus DNA. This chimera might be a very inter-
esting and very practicalmodelfor the future of
Despite these encouraging results, the vaccinia
vaccines, particularly for populations in develop-
format is no longer tenable because of potential
ing countries. Yellow fever chimera vaccines are
side effects, particularly in Africa, where HIV
already undergoing trials for Japanese Encephal-
itis and Dengue, so the idea is not new. However,
The population to be served is poor and the
these last two diseases are caused by flaviviruses,
virus is endemic in an area with poor or absent
and the chimeras replace the yellow fever E
infrastructure, so a Lassa virus vaccine must be
proteins with the proteins of the related virus of
cheap, easily and safely administered, and stable.
interest. A yellow fever/Lassa chimera requires a
Since we have now clearly shown that a recombi-
different approach – that of stable insertion of an
nant vaccine bearing the glycoprotein affords
protection, the first issue is the delivery formatfor this protein. There are severaloptions. Firstly anon-pathogenic vaccine such as Mopeia mightwork well, but safety concerns are likely to
complicate its use. The attraction of the live
attenuated vaccine is that a single shot, as with
The population at risk encompasses most of the
the 17D yellow fever vaccine, might well protect
population of West Africa to Nigeria, of the order
for life – or at least for a considerable number of
of 200 million or more by the time we have a
years – particularly for those living in endemic
vaccine in place. The people most at risk are the
areas and exposed to virus, who might therefore
poorest of the ruralinhabitants and localhealth
receive a silent, natural boost to immunity from
care workers, who should be the first people
time to time. Given the remoteness and mobility of
offered the vaccine. Since the epidemiological
the target population, this is a highly desirable
profile of Lassa fever is similar to that for
property for any vaccine candidate. However, live
yellow fever, long-term objectives should envisage
vaccines require a cold chain, and in the condi-
inclusion of a Lassa vaccine in EPI for West Africa.
tions where the vaccine is most needed, this
However, decisions such as these will depend
largely on the format of the final vaccine,
Killed vaccines have been shown to be quite
particularly the recommended schedule for boo-
ineffective, and are excluded from consideration.
Poxviruses, such as the Canarypox virus [45], (an
In view of the high risk to hospitalstaff in
avian virus which does not replicate in mam-
endemic areas, all medical care personnel in the
malian cells) is a possibility, but the question iswhether its immunogenicity will be sufficient, and
endemic area should be offered the vaccine.
how many doses might be needed to achieve
Recently aid workers, both UN personnel and
protection. These recombinants are also very
employees of non-governmental organizations
have been infected in West Africa, and all
A Lassa fever vaccine requires new approaches.
travellers to the region who expect to venture
DNA vaccines would offer much in the way of
outside the strict confines of the major cities and
low cost, stable vaccines, but several booster doses
the best hotels should be vaccinated. Finally,
might be needed. Another approach is to use the
laboratory staff in any country likely to receive
17D strain of yellow fever virus as a vehicle,
specimens from Lassa virus infected patients, staff
preferably without losing yellow fever anti-
caring for sick returning travellers from Lassa
genicity, since the population at risk for Lassa
endemic regions, or those working with Lassa
fever, is also exposed to yellow fever [46]. For the
virus in a research capacity, would also benefit
long term, a yellow fever/Lassa fever chimera
from the vaccine. Indeed the existence of the
vaccine for use in EPI in West Africa is a very
vaccine should allow the virus to be downgraded
attractive solution, however, this would require a
to BSL3, since at that point it would not only be
cold chain. The cDNA clone of yellow fever could
treatable but vaccine preventable. This would give
also be used as a vehicle for delivery of Lassa
enormous impetus to future research.
Copyright # 2001 John Wiley & Sons, Ltd.
Rev. Med. Virol. 2001; 11: 331–341.
the 1940s. These epidemics are the consequence of
failure of political will to implement those
The economic and political obstacles to producing
vaccines that already exist [47]. This can be
an effective Lassa fever vaccine far outweigh the
blamed on inertia, disinterest, corruption, civil
practicaland scientific problems. In the past two
strife; but the fact remains that the people needing
decades no vaccine destined primarily for popula-
the vaccines do not receive them, and for the most
tions in developing countries has been developed
part the global institutions such as the World
and marketed. The exception is the Candid 1
Health Organization or the UN seem relatively
vaccine for AHF, but it must be remembered that
this vaccine was developed and tested by the
There is some hope. Poor populations may lack
United States Army, whose priorities are rightly
education, but they are not unintelligent, and not
protection of military personnel. Even this vaccine
devoid of resources. Even the poorest will find a
is somewhat of a fortunate aberration since the
few pence for the welfare of his or her family if he
strategic importance of AHF is unclear. The fear
or she perceives the danger and appreciates the
of biological warfare, and manipulation of the
benefit of the commodity offered. Free vaccines
haemorrhagic fever viruses, including Lassa virus,
and medications often become devalued in the
to produce weapons has spurred some publicly
eyes of the recipient who has made no investment,
funded research. The fact remains, however,
however small, and who may not wholly under-
that pharmaceuticalcompanies, understandabl
stand or trust the motives of outsiders who come
answerable to their shareholders, do not and will
to give them. If the system is unreliable and
not consider a Lassa fever vaccine as a marketable
corrupt the mistrust is exacerbated. An effort to
vaccine. Public and private funding agencies may
market vaccines at a low price and to educate the
support some research, but not at the levels
population at the same time about the benefits is
needed to produce a human use vaccine for an
‘orphan’ disease. The costs and complexity of
Among the viralhaemorrhagic fevers, Lassa
conducting clinical trials continue to spiral out of
fever along with Hantaviruses afflicts the greatest
sight. The ethicalissues surrounding HIV vaccines
number of victims, and presents us with the
and their testing in the field are becoming
realface of an emerging disease that coul
increasingly contentious, and adding considerably
threaten larger communities. We have shown
to costs of trials. The truth today is that vaccines
that an effective and safe vaccine can be made
are developed for the wealthy countries where
and could be avidly pursued. The challenge to us
return on investment can be accurately predicted,
now is to overcome the political and economic
and their cost is rising dramatically. At the same
obstacles, and put this much-needed vaccine in
time the poorer countries that need the vaccines
are becoming relatively poorer, and the hope ofvaccines tailored for their needs diminishes.
Solutions to these problems are not easy to
1. Buckley SM, Casals J, Downs WG. Isolation and
imagine. Funding from a large charitable organi-
antigenic characterization of Lassa virus. Nature
sation might be adequate to launch the vaccine.
While the vaccine might fall under the orphan
2. Buckley SM, Casals J. Lassa fever, a new virus
biological development programme of the Food
disease of man from West Africa. 3. Isolation and
and Drug Administration, this is unlikely to be
characterization of the virus. Am J Trop Med Hyg
sufficient inducement for private development.
Testing, manufacturing and licensing the vaccine
3. Bishop DH. Arenaviridae and their replication. In
within the region present challenges which will
Fields Virology, 2nd edn, Fields BN, Knipe DM (eds). Raven Press: New York, 1993; 1231–1243.
not be solved until there is strong international
4. McCormick JB, Webb PA, Krebs JW, Johnson KM,
leadership, funding, and some difficult political
Smith ES. A prospective study of the epidemiology
decisions made. The problem does not end even
and ecology of Lassa fever. J Infect Dis 1987; 155:
then. In 2000 and 2001 we are again seeing large
epidemics of yellow fever, a haemorrhagic fever
for which we have had a safe, cheap vaccine since
analysis of the Arenaviridae: patterns of virus
Copyright # 2001 John Wiley & Sons, Ltd.
Rev. Med. Virol. 2001; 11: 331–341.
evolution and evidence for cospeciation between
21. Price ME, Fisher Hoch SP, Craven RB, McCormick
arenaviruses and their rodent hosts. Mol Phylogenet
JB. A prospective study of maternaland fetal
outcome in acute Lassa fever infection during
6. Bowen MD, Rollin PE, Ksiazek TG, et al. Genetic
pregnancy. Br Med J 1988; 297: 584–587.
diversity among Lassa virus strains. J Virol 2000; 74:
22. Cummins D, McCormick JB, Bennett D, et al. Acute
sensorineuraldeafness in Lassa fever [see com-
7. Wulff H, McIntosh BM, Hamner DB, Johnson KM.
ments]. JAMA 1990; 264: 2093–2096.
Isolation of an arenavirus closely related to Lassa
23. Bishop DH, Auperin DD. Arenavirus gene structure
virus from Mastomys natalensis in south-east Africa.
and organization. Curr Top Microbiol Immunol 1987;
Bull World Health Organ 1977; 55: 441–444.
8. Riviere Y. Mapping arenavirus genes causing viru-
24. Salvato MS, Schweighofer KJ, Burns J, Shimomaye
lence. Curr Top Microbiol Immunol 1987; 133: 59–65.
EM. Biochemicaland immunologicalevidence that
9. Djavani M, Lukashevich IS, Salvato MS. Sequence
the 11 kDa zinc-binding protein of lymphocytic
comparison of the large genomic RNA segments of
choriomeningitis virus is a structuralcomponent
two strains of lymphocytic choriomeningitis virus
of the virus. Virus Res 1992; 22: 185–198.
differing in pathogenic potentialfor guinea pigs.
25. Ruo SL, Mitchell SW, Kiley MP, Roumillat LF,
Fisher Hoch SP, McCormick JB. Antigenic related-
10. Fraser DW, Campbell CC, Monath TP, Goff PA,
ness between arenaviruses defined at the epitope
Gregg MB. Lassa fever in the Eastern Province of
level by monoclonal antibodies. J Gen Virol 1991; 72:
Sierra Leone, 1970–1972. I. Epidemiologic studies.
Am J Trop Med Hyg 1974; 23: 1131–1139.
26. Maiztegui JI, Fernandez NJ, de Damilano AJ.
11. Yalley Ogunro JE, Frame JD, Hanson AP. Endemic
Efficacy of immune plasma in treatment of Argen-
Lassa fever in Liberia. VI. Village serological sur-
tine haemorrhagic fever and association between
veys for evidence of Lassa virus activity in Lofa
treatment and a late neurological syndrome. Lancet
County, Liberia. Trans R Soc Trop Med Hyg 1984; 78:
27. Jahrling PB. Protection of Lassa virus-infected
12. Tomori O, Fabiyi A, Sorungbe A, Smith A,
guinea pigs with Lassa-immune plasma of guinea
McCormick JB. Viralhemorrhagic fever antibodies
pig, primate, and human origin. J Med Virol 1983; 12:
in Nigerian populations. Am J Trop Med Hyg 1988;
28. Johnson KM, McCormick JB, Webb PA, Smith ES,
13. Webb PA, McCormick JB, King IJ, et al. Lassa fever
Elliott LH, King IJ. Clinical virology of Lassa fever
in children in Sierra Leone, West Africa. Trans R Soc
in hospitalized patients. J Infect Dis 1987; 155:
14. Monson MH, Cole AK, Frame JD, Serwint JR,
29. Ter Meulen J, Badusche M, Kuhnt K, et al. Charact-
Alexander S, Jahrling PB. Pediatric Lassa fever: a
erization of human CD4(+) T-cell clones recogniz-
review of 33 Liberian cases. Am J Trop Med Hyg
ing conserved and variable epitopes of the lassa
virus nucleoprotein. J Virol 2000; 74: 2186–2192.
15. Fisher Hoch SP, Tomori O, Nasidi A, et al. Review of
30. Fisher Hoch SP, Price ME, Craven RB, et al. Safe
cases of nosocomialLassa fever in Nigeria: the high
intensive-care management of a severe case of Lassa
price of poor medicalpractice. Br Med J 1995; 311:
fever with simple barrier nursing techniques. Lancet
16. Monath TP, Mertens PE, Patton R, et al. A hospital
epidemic of Lassa fever in Zorzor, Liberia, March–
31. Fisher Hoch SP. Stringent precautions are not
April1972. Am J Trop Med Hyg 1973; 22: 773–779.
advisable when caring for patients with viral
17. Allan R, Ladbury R, Skinner K, Mardel S. Emer-
haemorrhagic fevers. Rev Med Virol 1993; 3: 7–13.
gence of epidemic Lassa fever during civilconflict
32. Helmick CG, Webb PA, Scribner CL, Krebs JW,
in Sierra Leone. International Conference on Emerging
McCormick JB. No evidence for increased risk of
Infectious Diseases 1998; 21.16;134.
Lassa fever infection in hospitalstaff. Lancet 1986; 2:
18. Lassa fever, imported case, Netherlands [news].
33. Maiztegui JI, McKee KT Jr, Barrera Oro JG, et al.
19. Lassa fever imported to England. Commun Dis Rep
Protective efficacy of a live attenuated vaccine
against Argentine hemorrhagic fever. AHF Study
20. McCormick JB, King IJ, Webb PA, et al. A case-
Group. J Infect Dis 1998; 177: 277–283.
controlstudy of the clinicaldiagnosis and course of
34. Kiley MP, Lange JV, Johnson KM. Protection of
Lassa fever. J Infect Dis 1987; 155: 445–455.
rhesus monkeys from Lassa virus by immunisation
Copyright # 2001 John Wiley & Sons, Ltd.
Rev. Med. Virol. 2001; 11: 331–341.
with closely related Arenavirus [letter]. Lancet 1979;
pigs from Lassa fever by vaccinia virus recombi-
nants expressing the nucleoprotein or the envelope
35. Fisher-Hoch SP, Hutwagner L, Brown B, McCormick
glycoproteins of Lassa virus. Virology 1989; 171:
JB. Effective vaccine for lassa fever. J Virol 2000; 74:
42. Auperin DD. Construction and evaluation of
36. Fisher Hoch SP, McCormick JB, Auperin DD, et al.
recombinant virus vaccines for Lassa fever. In The
Protection of rhesus monkeys from fatalLassa fever
Arenaviridae, Salvato MS (ed.). Plenum Press: New
by vaccination with a recombinant vaccinia virus
containing the Lassa virus glycoprotein gene. Proc
43. Morrison HG, Goldsmith CS, Regnery HL, Auperin
Natl Acad Sci U S A 1989; 86: 317–321.
DD. Simultaneous expression of the Lassa virus N
and GPC genes from a single recombinant vaccinia
inactivation of Lassa, Ebola, and Marburg viruses
virus. Virus Res 1991; 18: 231–241.
and effect on clinical laboratory analyses. J Clin
44. Auperin DD, McCormick JB. Nucleotide sequence
of the Lassa virus (Josiah strain) S genome RNA and
38. Clegg JC, Oram JD. Molecular cloning of Lassa virus
amino acid sequence comparison of the N and GPC
RNA: nucleotide sequence and expression of the
proteins to other arenaviruses. Virology 1989; 168:
nucleocapsid protein gene. Virology 1985; 144:
45. Tartaglia J, Excler JL, El Habib R, et al. Canarypox
39. Clegg JC, Lloyd G. Vaccinia recombinant expressing
virus-based vaccines: prime-boost strategies to
guinea pigs against Lassa fever. Lancet 1987; 2:
against HIV. AIDS Res Hum Retroviruses 1998; 14
40. Auperin DD, Esposito JJ, Lange JV, et al. Construc-
46. Rice CM, Lenches EM, Eddy SR, Shin SJ, Sheets RL,
tion of a recombinant vaccinia virus expressing the
Strauss JH. Nucleotide sequence of yellow fever
Lassa virus glycoprotein gene and protection of
virus: implications for flavivirus gene expression
guinea pigs from a lethal Lassa virus infection. Virus
and evolution. Science 1985; 229: 726–733.
47. Ruff TA. Immunisation strategies for viraldiseases
41. Morrison HG, Bauer SP, Lange JV, Esposito JJ,
in developing countries. Rev Med Virol 1999; 9:
McCormick JB, Auperin DD. Protection of guinea
Copyright # 2001 John Wiley & Sons, Ltd.
Rev. Med. Virol. 2001; 11: 331–341.
PHOTOREFRACTIVE EFFECT IN LITHIUM NIOBATE CRYSTALS OF VARIABLE COMPOSITIONS AND ITS MANIFESTATION IN RAMAN SPECTRA N.V. Sidorov1 P.G. Chufyrev1, M.N. Palatnikov1, V.M. Voskresensky1, and 1Institute of Chemistry and Technology of Rare Elements and Mineral Raw Materials KSC RAS, 2Institute of Solid State Physics, University of Latvia, 8 Kengaraga str., Riga, LV-1063, Latvia Results of Ra
Dental Care Guidelines Self-care Diabetes patients must maintain a rigorous oral self-care regimen to minimize oral health problems that may complicate glycemic control since the mouth is the first part of the digestive process. Regular visits (generally twice a year) to their dentist for a dental examination and dental prophylaxis and necessary radiographs are recommended to achieve an
 Rev. Med. Virol. 2001; 11: 331–341.
Rev. Med. Virol. 2001; 11: 331–341.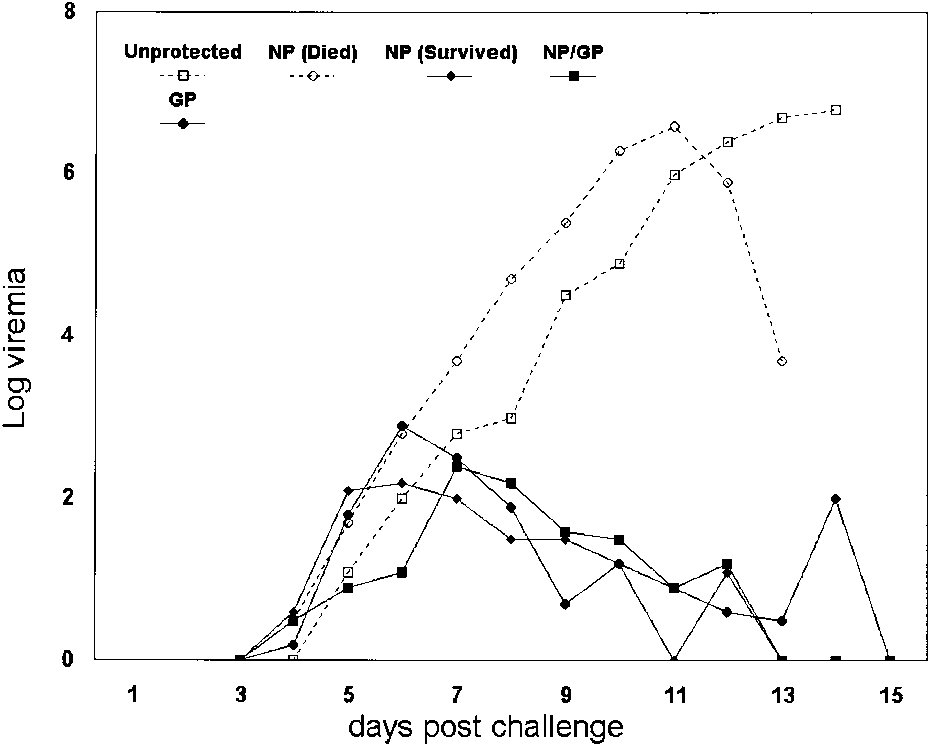 vaccinated with all proteins lived (90% survival).
vaccinated with all proteins lived (90% survival).