Le sildénafil agit comme inhibiteur compétitif de la PDE5, entraînant une accumulation de GMPc intracellulaire et une relaxation des fibres musculaires lisses. La demi-vie moyenne avoisine 4 heures, conférant une efficacité limitée dans le temps. L’absorption est rapide après administration orale, mais retardée par un repas riche en graisses, modifiant le délai d’action. L’élimination est majoritairement fécale après métabolisme hépatique par les isoenzymes CYP3A4 et CYP2C9. Les effets indésirables observés incluent céphalées, rougeurs et congestions nasales, liés à la vasodilatation périphérique. Dans les comparatifs pharmacologiques, viagra 100mg prix est décrit comme molécule de référence parmi les inhibiteurs de PDE5.
Eel_6420
Evolutionary Ecology Research, 2002, 4: 1017–1032 Generations of the polyphenic butterfly Araschnia levana differ in body design 1Department of Zoology, School of Biological Sciences, University of Southern Bohemia and2Institute of Entomology, Czech Academy of Sciences, Branisˇovská 31,CZ-370 05 Cˇeské Budeˇjovice, Czech RepublicABSTRACT
The European Map Butterfly, Araschnia levana (Nymphalidae), represents a classicalexample of seasonal polymorphism, in which summer and spring generations differ strikinglyin wing colour pattern. Although the physiological background to the formation of thetwo forms is well known, few studies have explored its possible ecological significance. Basedon previous findings that both population number and body size tend to be larger in summer,we examined the hypothesis that the summer generation is more capable of long-distancedispersal. We re-analysed mark–recapture data on the butterfly using the multi-strata modellingdesign in the program MARK, compared biomechanical design traits of the spring- andsummer-generation butterflies using canonical variate analysis, and tested for inter-generationdifferences in allometry relations of design traits to body size. The summer butterflies hadshorter residence times and a higher probability of movements among capture sites thanthe spring butterflies, indicating greater mobility. Regarding the design traits, adults ofsummer-generation butterflies had a heavier thorax, lower abdomen to body mass ratio,larger wing area, less pointed wings and lower wing loading. The differences in design traitsremained significant even after filtering out the effect of sex. Multiple regressions of the bio-mechanical traits against dry body mass confirmed the above differences between generationsand showed that the differences were not only due to different body size. The results supportthe notion of greater mobility of summer-generation A. levana and suggest the possibilitythat differential dispersal is much more widespread in butterflies with multiple generations. This is well known in insect groups that demonstrate obvious wing size polymor-phism, and might manifest itself in butterflies through more subtle shifts in body designproportions. Keywords: allometry, biomechanical design, canonical variate analysis, dispersal, Lepidoptera,seasonal polyphenism.
* Address all correspondence to Martin Konvic˘ka, Department of Zoology, School of Biological Sciences,University of Southern Bohemia, Branisˇovská 31, CZ-370 05, Cˇeské Budeˇjovice, Czech Republic. e-mail:[email protected] the copyright statement on the inside front cover for non-commercial copying policies. INTRODUCTION
The European Map Butterfly, Araschnia levana (Linnaeus, 1758), is a striking example ofseasonal polyphenism, the existence of seasonally different generations. Spring and summergenerations differ in wing colour patterns to such an extent that they were originally listedas different species (cf. Nijhout, 1991; Windig and Lammar, 1999). The formation of colourpattern is controlled by the timing of ecdysteroid release after pupation (Koch, 1992, 1996). The timing itself is controlled by length of day (short days induce diapause). Although thephysiological background to the formation of the two forms is well known, there isuncertainty regarding the ecological significance of the phenomenon. Windig (1999) andWindig and Lammar (1999) studied the evolutionary genetics of the species and foundthat various wing pattern traits reacted differently to environmental variation and did notoverlap across environments. They found neither individual differences in heritability ofwing-pattern traits, nor genetic correlations among the traits across different environments. Thus, the two forms were completely discrete, and the authors speculated on such poly-phenism as a step towards speciation. However, the possible adaptive value of having twogenerations that differ entirely in colour pattern is not understood in this species.
One of the hypotheses explaining seasonal polyphenism assumes that phenotypic differ-
ences in different generations allow a species to cope with seasonally changing environments(Shapiro, 1976; Brakefield, 1996). If true, then different selective forces should act ondifferent forms of A. levana. In a previous paper (Fric and Konvic˘ka, 2000), we comparedthe population structure and behaviour of the two generations and found that diurnalperiodicity and mate-locating behaviour were essentially identical in spring and summerbutterflies. However, the summer generation was more numerous, and we obtained indirectevidence of its greater mobility, since the summer generation showed a distinctly open(sensu Warren, 1992) population structure. The summer-generation butterflies are alsolarger (Windig, 1999); larger size is likely to be beneficial to dispersing butterflies, since itincreases their chances for successful colonization of new habitats (Kuussaari et al., 1996;Thomas et al., 1998). Thus, we hypothesized that the butterflies of the spring generationmight preferentially remain on sites where they developed from hibernating pupae, whereasthe summer butterflies might tend to leave their sites of emergence.
Here, we explore further the hypothesis that summer-generation A. levana are more
mobile. We first re-analyse the mark–recapture data of Fric and Konvic˘ka (2000) andcompare directly mobility-related parameters of the two generations. We use a modellingmethod that estimates the parameters of residence time (see below), capture probability andrate of migration between capture sites.
Second, we analyse differences in ‘biomechanical design’ between spring and summer
butterflies (Wickman, 1992): in allocation of body mass and in shape and area of wings. Differences among species that are exposed to differing selective forces were investigated inthe context of palatability and predation (Chai and Srygley, 1990; Marden and Chai, 1991;Srygley and Kingsolver, 1998; Kingsolver and Srygley, 2000) and in studies of mating tactics(Wickman, 1992). More relevantly, biomechanical traits may differ within individuals of thesame species originating from isolated and expanding populations (Thomas et al., 1998; Hillet al., 1999a,b; Van Dyck and Matthysen, 1999). Adult butterflies from recently expandingpopulations, as well as individuals from fragmented landscapes, tend to be larger, whichenhances their ability to colonize new habitats. They also tend to have relatively heavierthoraxes, due to a higher investment in flight muscles, and larger wing areas (Thomas et al.,
1998; Hill et al., 1999a); that translates into a lower wing loading (Roff and Fairbairn,1991). Windig (1999) and Windig and Lammar (1999) reported summer-generationA. levana to be larger (measured as adult wing area) but, to our knowledge, biomechanicaltraits have never been analysed across different generations in butterflies.
Third, we analyse patterns of allometric differences between adults of the two gener-
ations to exclude the possibility that differences in design are only due to the different sizesof the butterflies. Modelling mobility parameters
The mark–recapture data were obtained during 1997 using a standard protocol. We markedbutterflies within the study site, which was a system of wet meadows and deciduouswoodlots, for entire flight periods of the spring (5–26 May) and summer (16 July–13August) generations, and recorded sex and position of each capture in four separate sectionsof the study site (details in Fric and Konvic˘ka, 2000).
We used the program MARK (White and Burnham, 1999; Cooch and White, 2001) to
analyse the data. The program considers pre-defined individual covariates of markedanimals (such as sex, site, time of capture, etc.) and allows the construction of multiplemodels that vary in their numbers of parameters. The parameters are computed by a gener-alized linear modelling procedure (Lebreton et al., 1992); their parsimony is compared usingthe quasi-likelihood AIC values (QAICc) (Franklin, 1999).
Since we had four possible locations of capture of each individual, we applied the
multi-strata design with the logit link function (Brownie et al., 1993). The capture sites ofindividual butterflies were entered into the data matrix with sex as the individual covariate. The design allowed us to consider movements of the butterflies among the capture sitesand to estimate the following parameters: relative residence (i.e. the probability that abutterfly remains alive at the site of its original capture, S); capture probability (p); and theprobability of movement between capture points (Ψ) (Cooch and White, 2001).
The program selects the model that provides the most parsimonious explanation of
the mark–recapture data. However, if the ‘best’ models for different data sets differ intheir constituent parameters, their parameter values cannot be compared directly. Since thiswas the case, we looked for a pair of simpler models that contained mutually matchingparameters and that still explained a sufficiently high proportion of the original deviance(Lebreton et al., 1992). We used these ‘simplified’ models to compare their parameters. Biomechanical design
In 1997 and 2001, we collected males and females of both generations of A. levana (springgeneration: 22ΗΗ, 13ΘΘ; summer generation: 36ΗΗ, 20ΘΘ) in the field. The materialoriginated from four sites, all of them within 25 km2, in the close vicinity of Cˇeské Budeˇjovice,South Bohemia, Czech Republic (49Њ02ЈN, 14Њ30ЈE). We processed the butterflies as inWickman (1992); that is, we weighed their fresh mass, dried them at 60ЊC to constant weight,and weighed their total dry mass, thorax mass and abdomen mass. We placed the wings ina natural position as in flight and digitized their images. We measured the length of theforewings, wing area and the orthogonal projection of the centroid of the wing area on wing
diameter (hereafter ‘centroid projection’), using the program Scion Image Beta 3b (ScionCorp., 1998). We used the above values, hereafter referred to as ‘raw’ variables, to compute thefollowing ‘derived’ variables: (1) flight muscle ratio, i.e. thorax mass to total body mass; (2)abdomen ratio, i.e. abdomen mass to total body mass; (3) wing loading, i.e. fresh body massdivided by wing area; (4) aspect ratio, i.e. wing span squared divided by wing area; and (5)radius of the first moment of wing area, i.e. the spanwise position of the normalized chordthrough the centroid of the wing area as a fraction of the wing length (hereafter ‘radius’). Thelatter variable is a measure of the pointedness of wings (Wickman, 1992).
We thus had four classes of butterflies – spring males, spring females, summer males
and summer females – and our main concern was whether the classes differed in theirbiomechanical traits. In other words, we examined the hypothesis that the biomechanicaltraits, being the explanatory variables, can be used to separate the classes. We used canonicalvariate analysis (CVA), a variant of classical linear discrimination analysis, and performedthree separate analyses. The first analysis included all possible explanatory variables; thesecond included only the raw variables and the third only the derived variables. We usedCANOCO for Windows, version 4.0 (Ter Braak and Sˇmilauer, 1998), which computes CVAas a variant of canonical correspondence analysis with Hill’s scaling and handles thediscriminated classes as dummy ‘species data’. We first tested individual effects of allpotential explanatory variables, and then forward-selected for the best sets of explanatoryvariables, using CANOCO’s variance inflation factors for eliminating redundant variables. We assessed the statistical significance of the results using non-parametric Monte Carlopermutation tests (MCPT; 999 runs per analysis).
To eliminate the possibility that significant differentiation among the classes was due to
differences between males and females, we subsequently repeated the analyses with twoclasses only, spring and summer butterflies. Sex was included as a categorical covariablewhose influence on the statistical significance of the results was filtered out (Ter Braak andSˇmilauer, 1998).
We also controlled for the effects of year and site, again using CVA, with the classes as
dummy species data and year/site as categorical explanatory variables. Allometry
Differences in biomechanical design between generations and between the sexes couldbe caused solely by different allometric growth patterns in individuals of different size(Klingenberg, 1996). To exclude this possibility, we constructed multiple-regression modelsin which the effects tested were the design variables from the previous analyses and theexplanatory variables were the dry mass of individuals, sex, generation (the two as categor-ical predictors) and all possible interactions. Dry mass was included in the models asa surrogate of body mass to force the size-related variation to the relationships, whilethe inclusion of the two categorical predictors tested for differences due to generation orsex independent of body size. The significant relationships explained by the categoricalpredictors indicate different heights of the respective regression lines; the relationshipsexplained by interactions indicate differences in regression slopes.
Possible part–whole bias (in the models with the ‘derived variables’, such as the flight
muscle ratio) was eliminated because total dry mass was included in the multiple regres-sions. Hence the variation due to the categorical predictors and interactions was assessedafter accounting for the background variation.
All the regressions were constructed as linear models, using the program S-plus
(S-Plus 2000, 1999). To avoid the false rejection of the null hypotheses in running multiplemodels, we used the Bonferroni correction, setting α to 0.005 (ten multiple-regressionmodels). Modelling mobility parameters
In total, we caught 445 spring-generation butterflies (320ΗΗ, 125ΘΘ) and 960 summer-generation butterflies (759ΗΗ, 201ΘΘ). The two ‘best’ models (for spring and summergenerations) selected by the MARK program differed substantially in their number ofconstituent parameters (Table 1). In the spring generation, the residence of the butterflieswas independent of sex and position of capture, and males had a higher capture probability
Table 1. The estimated parameter values for the best models that fitted the mark–release–recapture data for the spring and summer generations of Araschnia levana Spring generation S (.) Summer generation S (A ΗΗ) Note: S = residence, p = capture probability, Ψ = probability of movement. Table 2. Evaluation of the best models fitting the mark–release–recapture data of the spring and summer generations of Araschnia levana (‘Best’), and comparison with models that contained identical parameters as the best model for the other generation (‘Equivalent’), with the full model that contained all possible parameters, and with the most reduced models (‘Simplified’) Spring generation Best: S(.) p(sex) Ψ (A = B, rest: every-to-all) Equivalent: S(site*sex) p(site*sex) Ψ(.)
Simplified model: S(.) p(.) Ψ(.)
Summer generation Best: S(site*sex) p(site*sex) Ψ (.) Equivalent: S(.) p(sex) Ψ(A = B, rest: every-to-all)
Simplified model: S(.) p(.) Ψ(.)
Note: S = residence, p = capture probability, Ψ = probability of movement.
than females. The probability of movement between capture sites was high for two sitesreferred to as A and B in Fric and Konvic˘ka (2000) and very low for the other sites (Table 1).
The best model for the summer generation contained 17 parameters (Table 2). Both
residence values and the probability of capture differed between the sexes and among sites,whereas the probability of moving between capture sites did not differ for individual sites. The residence values were lower than in the spring generation for both sexes at most of thesections. An exception was the residence of females at site B, where we regularly observedthem to lay eggs at a large patch of Urtica dioica, the host plant of the butterfly.
It was not possible to compare the parameters of the two best models directly. The only
simplified models that contained identical parameters for both generations, and at the sametime achieved sufficiently low deviance values, were the most reduced models. They assumedthat residence time, capture probability and probability of moving between sites wereidentical for both sexes and all capture sections (Table 2). The parameter values of thesesimplified models suggested, first, that the spring butterflies had longer residence times(S = 0.654, standard error = 0.187) than the summer butterflies (S = 0.405, standarderror = 0.028); second, capture probability was higher in summer (spring: p = 0.121,standard error = 0.062; summer: p = 0.285, standard error = 0.035); and third, theprobability of moving between capture sites was higher in the summer generation (spring:Ψ = 0.092, standard error = 0.039; summer: Ψ = 0.150, standard error = 0.015).
Hence, the shorter residence times of summer butterflies supported our previous sugges-
tion of higher dispersal capability in the summer generation. Biomechanical design
Neither year nor site of origin of the material influenced the biomechanical design of thebutterflies. (CVA, year: F = 1.18, P = 0.30; site: 1st axis F = 0.33, P = 0.99; all axes F = 0.12,
P = 0.99). This allowed us to pool the samples from different years and sites in furtheranalyses.
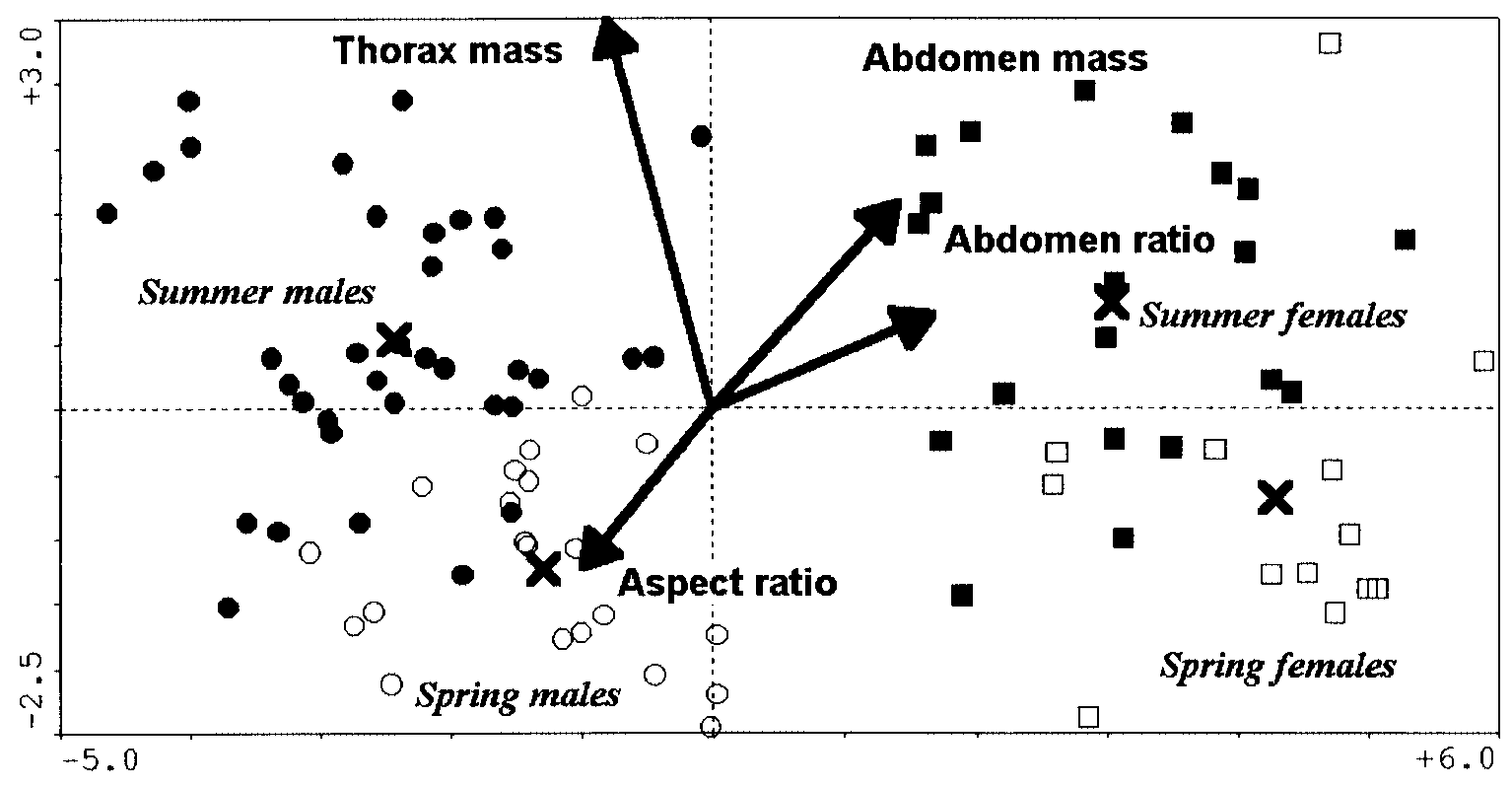
The CVA analysis with all variables convincingly and highly significantly separated the
four classes (Fig. 1, Tables 3 and 4). Butterflies of the summer generation were significantlylarger and had heavier thoraxes. In contrast, the butterflies of the spring generation hada larger wing loading (especially the females) and aspect ratio (especially the males). Thisindicates the capability of highly energetic, but costly flight. The discrimination using theraw variables was again highly significant (Fig. 2). Thus, the heavier thoraxes, heavierabdomens and larger wings of the summer generation hold even in absolute terms. Finally,the significant model that was based solely on derived variables showed that the effect ofabdomen ratio (higher in females) differentiated the sexes, whereas wing loading (higher inspring) separated the generations.
Filtering out the effect of sex on variation in the biomechanical data resulted in a
decrease in the variation explained by the individual models (Table 4). Still, the generalpattern remained preserved (Table 3). Summer-generation butterflies had significantlylarger and longer wings, heavier thoraxes and higher thorax muscle ratios. Springgeneration-butterflies had a heavier wing loading (Fig. 3).
In summary, the results suggest two trends. The first was a difference in design between
males and females: males were smaller than females in both generations, and had lighterabdomens and heavier thoraxes in both absolute and relative terms. They also had a higheraspect ratio, which is commonly associated with better flight manoeuvrability (Danforth,1989; Chai and Srygley, 1990). The second trend concerned the differences betweengenerations. The summer butterflies were larger in absolute terms and displayed a higherthorax muscle ratio (or a lower abdomen ratio, as the two measurements were negatively
Fig. 1. Canonical variate analysis of biomechanical traits of Araschnia levana. Raw and derived explanatory variables combined, sexes within generations treated separately. Only the variables selected by forward selection and included in the final model are shown. The scaling of darts for explanatory variables was multiplied by a factor of 5 for clarity. ᭺, spring males; ᮀ, spring females;
᭹, summer males; , summer females. × = centroids for individual classes. Table 3. Canonical variate analyses of biomechanical traits in males and females of spring- and summer-generation Araschnia levana: separate effects of single terms Note: Significance of effects of the terms was tested by Monte-Carlo permutation tests (999 permutations). The terms that achieved significance at P < 0.05 and had a variance inflation factor < 30 were entered into theconstruction of the final models.
# Denotes the variables with a variance inflation factor >30, which were thus excluded from the final models. *P < 0.05; **P < 0.01; ***P < 0.001. Table 4. Canonical variate analyses of biomechanical traits in males and females of spring- and summer-generation Araschnia levana: tests of final models Note: Significance of effects of the terms was tested by Monte-Carlo permutation tests (999 permutations). See Table 3 for the tests of model terms. *P < 0.05; **P < 0.01; ***P < 0.001.
correlated; R2 = −0.94 and P < 0.0001) and a lower wing loading. Since the CVA discrim-inations according to these traits remained highly significant when the variability attribut-able to sexual differences was filtered out, it follows that there were genuine inter-generationdifferences in biomechanical design. Fig. 2. Canonical variate analysis of biomechanical traits of Araschnia levana. Raw variables, sexes within generations treated separately. Only the variables selected by forward selection and included in the final model are shown. The scaling of darts for explanatory variables was multiplied by a factor of 5 for clarity. ᭺, spring males; ᮀ, spring females; ᭹, summer males; , summer females. × = centroids for individual classes. Allometry
The multiple regression models of the biomechanics variables against dry body mass, sexand generation and their interactions (Table 5) confirmed the larger investment by thespring generation in abdomen mass (also manifested as a higher wing loading) and thelarger investment by the summer generation in wing size and flight muscles.
Tests for the effects of generation were significant, after subtracting the trivial (and
highly significant) variation due to overall body size, for all the variables tested except thecentroid projection and the aspect ratio. That the factor ‘generation’ produced significantdifferences in the slope of the regression lines for eight of the ten models suggests that thebetween-generation patterns were not due to differences in body size alone. DISCUSSION
Both the comparisons of the residence and mobility parameters of the mark–recapturemodels, and the analyses of biomechanical traits, provided strong support for our initialhypothesis of greater mobility of summer-generation Araschnia levana butterflies. Thesummer-generation butterflies had shorter residence times (which, in fact, combines
Fig. 3. Canonical variate analysis of biomechanical traits of Araschnia levana. Derived variables, sexes within generations treated as covariables. Only the variables selected by forward selection are shown. The scaling of the dart for the explanatory variables was multiplied by a factor of 2 for clarity. ᭺, spring males; ᮀ, spring females; ᭹, summer males; , summer females; ᭡, centroid for the spring generation; ᭝, centroid for the summer generation.
survival and site-tenacity) and higher ‘emigration probability’ than the spring butterflies. However, the modelling approach provides only indirect insight into mobility. Nevertheless,it is one of the few approaches available for inter-generation comparison of the mobilityof this species. Alternatives, such as mark–recapture across an array of multiple habitatpatches (e.g. Hanski et al., 2000; Roland et al., 2000), would be technically prohibitive inA. levana because of its distinctly open population structure in summer.
The second line of evidence is the inter-generation differences in biomechanical design.
The summer generation conformed to what was expected for butterflies designed forcrossing relatively large distances. Not only were they larger, but they also had higher thoraxmuscle ratios and larger (in terms of both area and length) and less pointed wings. As aresult, their wing loading was lower than that of spring butterflies.
Thus, the differences between the spring and summer generations of A. levana were
congruent with those found in the satyrid butterfly Pararge aegeria (Berwaerts et al., 1998)and the lycaenid butterfly Plebejus argus (Hill et al., 1999a) from fragmented habitats, asopposed to continuous habitats. The patterns in the summer-generation A. levana corre-
−583.052 −24.4323 −1.4403 −0.9534 −0.3051
−28.2158 −1.0956 −0.4334 −4.0825 −0.0009 −25.5260 −28.2074
sponded similarly with the changes in biomechanical design reported for two butterflies,Pararge aegeria and Hesperia comma, during expansion of their ranges (Thomas et al.,1998; Hill et al., 1999b). Last but not least, the allometry analysis confirmed that theinter-generation differences in design were independent of differences in body size.
In contrast to our study, the studies cited in the previous paragraph used captive-reared
butterflies, which allowed family effects to be assessed – that is, the heritable components ofvariation in the biomechanical data. We did not study heritability of the design traits, sincewe were primarily interested in differences between generations. To obtain a captive lineageof A. levana spanning multiple generations would require rearing the species for at least2 years. This is not feasible at the moment, since the butterfly displays complex pre-matingbehaviour, including lekking by males (Fric and Konvic˘ka, 2000), and such inseminationtechniques as hand-pairing are not available. Also, working with captive-reared butterflies,the authors that studied Pararge aegeria, Plebejus argus and Hesperia comma worked withindividuals of the same imaginal age. This was not the case in our study, which could haveintroduced some ‘noisy’ variation into our data. For instance, the abdomen mass of femalesdecreases with age in butterflies, as progressively more eggs are laid. Despite this, the highlysignificant differences between generations found in our canonical variate analyses, and thefinding that the inter-generation differences were not caused solely by the effects of sex ortotal body size, render our main conclusion robust.
It can be argued that some of the design differences may not be connected to dispersal
capability. First, an increased flight muscle ratio, which we found in summer butterflies, maybe beneficial in reproductive competition. We have shown previously that the summergeneration is more numerous (Fric and Konvic˘ka, 2000) and reproductive competition mayintensify at high population densities. However, the summer-generation males had a lowerwing loading than the spring males. An increased wing loading translates into bettermanoeuvrability, which is of special concern for butterflies with a perching mate-locatingtactic (Wickman, 1992), a tactic used by males of both generations of A. levana (Fric andKonvic˘ka, 2000). This rules out the higher flight muscle ratio in summer males being a traitselected for by reproductive competition. Otherwise, the wing loading should have beenhigher in summer males. Also, an increase in flight muscle ratio for reproductive reasonsdoes not make sense in females, who do not depend on energetic flight for mate acquisition(cf. Wickman, 1992; Van Dyck et al., 1997).
The second trait closely connected to reproduction is relative abdomen mass. Abdomens
are filled with developing eggs in butterfly females and we agree with Rutowski (1997) thatthe larger absolute size of females is reproductively beneficial. However, both the absoluteand relative mass of the abdomen was higher in spring even after subtracting the effect ofbody size (Table 5). This suggests that summer-generation butterflies invest more resourcesin body structures other than the reproductive ones. This supports the finding of Dixon andKindlmann (1999), who observed that dispersing (winged) generations of aphids investrelatively less in the gonads than do sedentary (unwinged) generations. Although itis important for dispersing animals (whether aphids or butterflies) to invest heavily inreproduction – otherwise they would not establish their progeny at a new site – dispersalapparatus is costly, and building it diminishes investment in reproductive structures via atrade-off.
In summary, the evidence indicates that the spring generation of A. levana is more
sedentary and invests heavily in reproduction, whereas the summer generation tends todisperse in search of new inhabitable space. Such a strategy is perfectly reasonable: the
butterfly probably suffers increased mortality during winter diapause (cf. Windig, 1999),and any site in which the overwintering butterflies finish their development is thus bydefinition both relatively empty and suitable for reproduction. In summer, on the otherhand, population densities are much higher and it may pay for the butterflies to leave,exactly as proposed for mobile butterflies by Baker (1984).
This reflects the situation found in other insect groups, in which different generations are
polymorphic with respect to dispersal. Examples include aphids, Homoptera, Heteropteraand Orthoptera, in which sedentary generations typically have shortened or absent wings(Harrison, 1980; Waloff, 1980; Roff and Fairbarn, 1991; Rankin and Burchsted, 1992;Dixon and Kindlmann, 1999). Because no such dramatic dispersal-related adaptations arefound in butterflies, the possibility that different generations might vary in their mobilitywas never sought in this group, with the important exception of ‘classical’ migrants (e.g. Urquhart, 1960; Dingle et al., 1999). However, many butterflies that are not ‘classical’migrants still possess considerable dispersal abilities (Vandewoestijne et al., 1999), anddifferent abundances of seasonal broods have been observed in several species (e.g. Ohsaki,1980; Tabashnik, 1980; Pollard et al., 1997; Rothery and Roy, 2001). Good dispersalcapability and variation in abundance among generations are obvious prerequisites forinter-generational dispersal polymorphism. It follows that inter-generational differencesin dispersal capabilities might be more common in butterflies than previously thought. As exemplified by ‘hidden’ alary polymorphism in the Homoptera (Waloff, 1980), suchdifferences may not manifest themselves in different development of the wings, but byrelatively subtle shifts in body proportions. Also, and unlike in A. levana, inter-generationalpolymorphism in dispersal capability does not have to be associated with polyphenism inwing pattern and coloration.
We propose that differential dispersal should be beneficial for species that inhabit
seasonally changing environments, allowing individual generations to utilize varyingresources. If so, the patterns in biomechanical design of individual generations ofmultivoltine butterflies should be ‘ecologically predictable’. Specifically, the generationthat utilizes seasonally predictable but spatially unpredictable increases in resourcesshould be designed for better mobility. Examples of such situations should be sought, forexample, among temperate-zone Pieridae, which track ample supplies of their host plantsat cultivated fields in late summer, or among wetland-inhabiting temperate species,which should be more mobile during more humid months. The same might apply totropical species with dry-season and wet-season forms (cf. Windig et al., 1994; Dingle et al.,1999).
To test the above hypotheses, two lines of evidence should be explored. First, corre-
sponding differences in biomechanical design as those demonstrated for A. levana shouldbe found in a broader array of multivoltine species, including species that are notpolyphenic in wing patterns. Indeed, seasonal differences in ‘wing shape’ have been reportedfor an African polyphenic Precis butterfly (McLeod, 1984) and for the satyrid Parargeaegeria, in which the generations differ even in their behaviour (Van Dyck et al., 1997). Second, since biomechanics provides indirect evidence only for mobility, it should bedemonstrated that the generations that potentially differ in mobility do in fact utilize differ-ent ranges of habitats (or differ seasonally in abundances in particular habitat types). Inany case, studies of inter-generational differences in biomechanical traits appear to be acost-efficient way of revealing ecologically relevant patterns in dispersal and colonizationcapabilities of butterflies and other insect groups. ACKNOWLEDGEMENT
We are grateful to Oldrˇich Cˇíz˘ek for help with the material, to Jirˇí Benesˇ, Ferenc Kassai, PavelKindlmann, Jan Lepsˇ, Vojteˇch Novotný, Petr Sˇmilauer and Josef Settele for inspiring discussions onvarious drafts of the manuscript, and to Derek Roff for valuable comments and patient editing. Thestudy was funded by grant FRVSˇ 21-1295 to Z. Fric. REFERENCES
Baker, R.R. 1984. The dilemma: when and how to go or stay. In Biology of Butterflies, Symposium ofthe Royal Entomological Society of London, No. 11 (R.I. Vane-Wright and P.R. Ackery, eds),pp. 279–296. London: Academic Press.
Berwaerts, K., Van Dyck, H., Van Dongen, S. and Matthysen, E. 1998. Morphological and genetic
variation in the speckled wood butterfly (Pararge aegeria L.) among differently fragmented landscapes. Neth. J. Zool., 48: 241–253.
Brakefield, P.M. 1996. Seasonal polyphenism of butterflies and natural selection. Trends Ecol.Evol., 11: 275–277.
Brownie, C., Hines, J.E., Nichols, J.D., Pollock, K.H. and Hestbeck, J.B. 1993. Capture–
recapture studies for multiple strata including non-Markovian transitions. Biometrics, 49: 1173–1187.
Chai, P. and Srygley, R.B. 1990. Predation and the flight, morphology, and temperature of
neotropical rain-forest butterflies. Am. Nat., 135: 748–765.
Cooch, E. and White, G. 2001. Program Mark: Analysis of Data from Marked Individuals – ‘A GentleIntroduction’. Online at: http://canuck.dnr.cornell.edu/mark/
Danforth, B.N. 1989. The evolution of hymenopteran wings: the importance of size. J. Zool., 218:
Dingle, H., Zalucki, M.P. and Rochester, W.A. 1999. Season-specific directional movement in
migratory Australian butterflies. Aust. J. Entomol., 38: 323–329.
Dixon, A.F.G. and Kindlmann, P. 1999. Population dynamics of aphids. In Insect Populations
(J.P. Dempster and J.I.F.C. McLean, eds), pp. 207–230. Dordrecht: Kluwer Academic.
Dixon, A.F.G. and Kindlmann, P. 1999. Cost of flight apparatus and optimum body size of aphid
migrants. Ecology, 80: 1678–1690.
Franklin, A.B. 1999. Exploring Ecological Relationships in Survival and Estimating Rates ofPopulation Change Using Program MARK. Online at: http://www.cnr.colostate.edu/class_info/fw663/Mark.html
Fric, Z. and Konvic˘ka, M. 2000. Adult population structure and behaviour of two seasonal
generations of the European Map Butterfly, Araschnia levana, species with seasonal polyphenism (Nymphalidae). Nota Lepid., 23: 2–25.
Hanski, I., Alho, J. and Moilanen, A. 2000. Estimating the parameters of survival and migration of
individuals in metapopulations. Ecology, 81: 239–251.
Harrison, R.G. 1980. Dispersal polymorphism in insects. Annu. Rev. Ecol. Syst, 11: 95–118. Hill, J.K., Thomas, C.D. and Blakeley, D.S. 1999a. Evolution of flight morphology in a butterfly that
has recently expanded its geographic range. Oecologia, 121: 165–170.
Hill, J.K., Thomas, C.D. and Lewis, O.T. 1999b. Flight morphology in fragmented populations of
a rare British butterfly, Hesperia comma. Biol. Conserv., 87: 277–283.
Kingsolver, J.G. and Srygley, R.B. 2000. Experimental analysis of body size, flight and survival
in Pierid butterflies. Evol. Ecol. Res., 2: 593–612.
Klingenberg, C.P. 1996. Multivariate allometry. In Advances in Morphometrics (L.F. Marcus,
M. Corti, A. Loy, G.J.P. Naylor and D.E. Slice, eds), pp. 23–49. New York: Plenum Press.
Koch, P.B. 1992. Seasonal polyphenism in butterflies: a hormonally controlled phenomenon of
pattern formation. Zool. Jb. Physiol., 1996: 227–240.
Koch, P.B. 1996. Preadult changes of ecdysteroid and juvenile hormone titres in relation
to diapause and pigmental variations in two lepidopteran species, Cerura vinula and Araschnia levana (Lepidoptera: Notodontidae, Nymphalidae). Entomol. Gen., 20: 143–155.
Kuussaari, M., Nieminen, M. and Hanski, I. 1996. An experimental study of migration in the
glanville fritillary butterfly Melitaea cinxia. J. Anim. Ecol., 65: 791–801.
Lebreton, J., Burnham, K.P., Clobert, J. and Anderson, D.R. 1992. Modelling survival and testing
biological hypotheses using marked animals: a unified approach with case studies. Ecol. Monogr., 62: 67–118.
Marden, J.H. and Chai, P. 1991. Aerial predation and butterfly design – how palatability, mimicry,
and the need for evasive flight constrain mass allocation. Am. Nat., 138: 15–36.
McLeod, L. 1984. Seasonal polyphenism in African Precis butterflies. In The Biology of Butterflies
(R.I. Vane-Wright and P.R. Ackery, eds), pp. 313–315. London: Academic Press.
Nijhout, H.F. 1991. The Development and Evolution of Butterfly Wing Pattern. Washington, DC:
Ohsaki, N. 1980. Comparative population studies of three Pieris butterflies, P. rapae, P. melete and
P. napi, living in the same area. II. Utilization of patchy habitats by adults through migratory and non-migratory movements. Res. Popul. Ecol., 22: 163–183.
Pollard, E.H., Greatorex-Davies, J.N. and Thomas, J.A. 1997. Drought reduces breeding success of
the butterfly Aglais urticae. Ecol. Entomol., 22: 315–318.
Rankin, M.A. and Burchsted, J.C.A. 1992. The cost of migration in insects. Annu. Rev. Entomol., 37:
Roff, D.A. and Fairbairn, D.J. 1991. Wing dimorphisms and the evolution of migratory
polymorphisms among the Insecta. Am. Zool., 31: 243–251.
Roland, J., Keyghobadi, N. and Fownes, S. 2000. Alpine Parnassius butterfly dispersal: effects of
landscape and population size. Ecology, 81: 1642–1653.
Rothery, P. and Roy, D.B. 2001. Application of generalized additive models to butterfly transect
count data. J. Appl. Stat., 28: 897–909.
Rutowski, R.L. 1997. Sexual dimorphism and mating systems in butterflies. In The Evolutionof Mating Systems in Insects and Arachnids (J.C. Choe and B.J. Crespi, eds), pp. 257–272. Cambridge: Cambridge University Press.
1999. Guide to Statistics, Vol. 1. Seattle, WA: MathSoft, Data Analysis Products
Shapiro, A.M. 1976. Seasonal polyphenism. Evol. Biol., 9: 259–333. Srygley, R.B. and Kingsolver, J.G. 1998. Red-Wing Blackbird reproductive-behavior and the
palatability, flight performance, and morphology of temperate Pierid butterflies (Colias, Pieris, and Pontia). Biol. J. Linn. Soc., 64: 41–55.
Tabashnik, B.E. 1980. Population structure of Pierid butterflies. III. Pest population of Coliasphilodyce eriphyle. Oecologia, 47: 175–183.
Ter Braak, C.J.F. and Sˇmilauer, P. 1998. CANOCO Reference Manual and User’s Guide to Canocofor Windows. Software for Canonical Community Ordination (version 4). Wageningen: Centrefor Biometry Wageningen/Ithaca, NY: Microcomputer Power.
Thomas, C.D., Hill, J.K. and Lewis, O.T. 1998. Evolutionary consequences of habitat fragmentation
in a localised butterfly. J. Anim. Ecol., 67: 485–497.
Urquhart, F.A. 1960. The Monarch Butterfly. Toronto: University of Toronto Press. Van Dyck, H. and Matthysen, E. 1999. Habitat fragmentation and insect flight – a changing design
in a changing landscape. Trends Ecol. Evol., 14: 172–174.
Van Dyck, H., Matthysen, E. and Dhondt, A.A. 1997. Mate-locating strategies are related to relative
body length and wing color in the Speckled Wood Butterfly Pararge aegeria. Ecol. Entomol., 22: 116–120.
Vandewoestijne, S., Neve, G. and Baguette, M. 1999. Spatial and temporal population genetic
structure of the butterfly Aglais urticae L. (Lepidoptera, Nymphalidae). Mol. Ecol., 8: 1539–1543.
Waloff, N. 1980. Studies in grasslands leafhopeers (Auchenorrhyncha, Hopotera) and their natural
enemies. Adv. Ecol. Res., 2: 81–215.
Warren, M.S. 1992. Butterfly populations. In The Ecology of Butterflies in Britain (R.L.H. Dennis,
ed.), pp. 73–92. Oxford: Oxford University Press.
White, G.C. and Burnham, K.P. 1999. Program Mark: survival estimation from populations of
marked animals. Bird Study, 46 (suppl.): 120–138.
Wickman, P.O. 1992. Sexual selection and butterfly design – a comparative study. Evolution, 46:
Windig, J.J. 1999. Trade-offs between melanization, development time, and adult size in Inachis io
and Araschnia levana (Lepidoptera: Nymphalidae)? Heredity, 82: 57–68.
Windig, J.J. and Lammar, P. 1999. Evolutionary genetics of seasonal polyphenism in the map
butterfly Araschnia levana (Nymphalidae: Lepidoptera). Evol. Ecol. Res., 1: 875–894.
Windig J.J., Brakefield P.M., Reitsma, N. and Wilson, J.G.M. 1994. Seasonal polyphenism in
the wild: survey of wing patterns in five species of Bicyclus in Malawi. Ecol. Entomol., 19: 285–298.
Zar J.H. 1996. Biostatistical Analysis, 3rd edn. London: Prentice-Hall.
Last updated August 2011/ Due for review December 2013Poppers are a nitrite-based drug. Amyl nitrite is used medically to ease thechest pain caused by The drug gets its name from the small glasscapsules containing amyl nitrite for the treatment of angina which used tobe 'popped' under the nose and inhaled. Amyl and butyl nitrite started to beAs with any drug it may be wise to consider how u
azuma is offering a new afternoon tea service, including wonderful high tea menus & savoury options. this service is available monday to friday, 2pm–5pm. we hope you enjoy your azuma high tea experience. kyoto high tea – only $25 a savoury & sweet bento box served with your choice of tea or coffee sushi - salad rolls with asparagus, avocado, seasonal fruit salad with champag
 P = 0.99). This allowed us to pool the samples from different years and sites in furtheranalyses.
P = 0.99). This allowed us to pool the samples from different years and sites in furtheranalyses.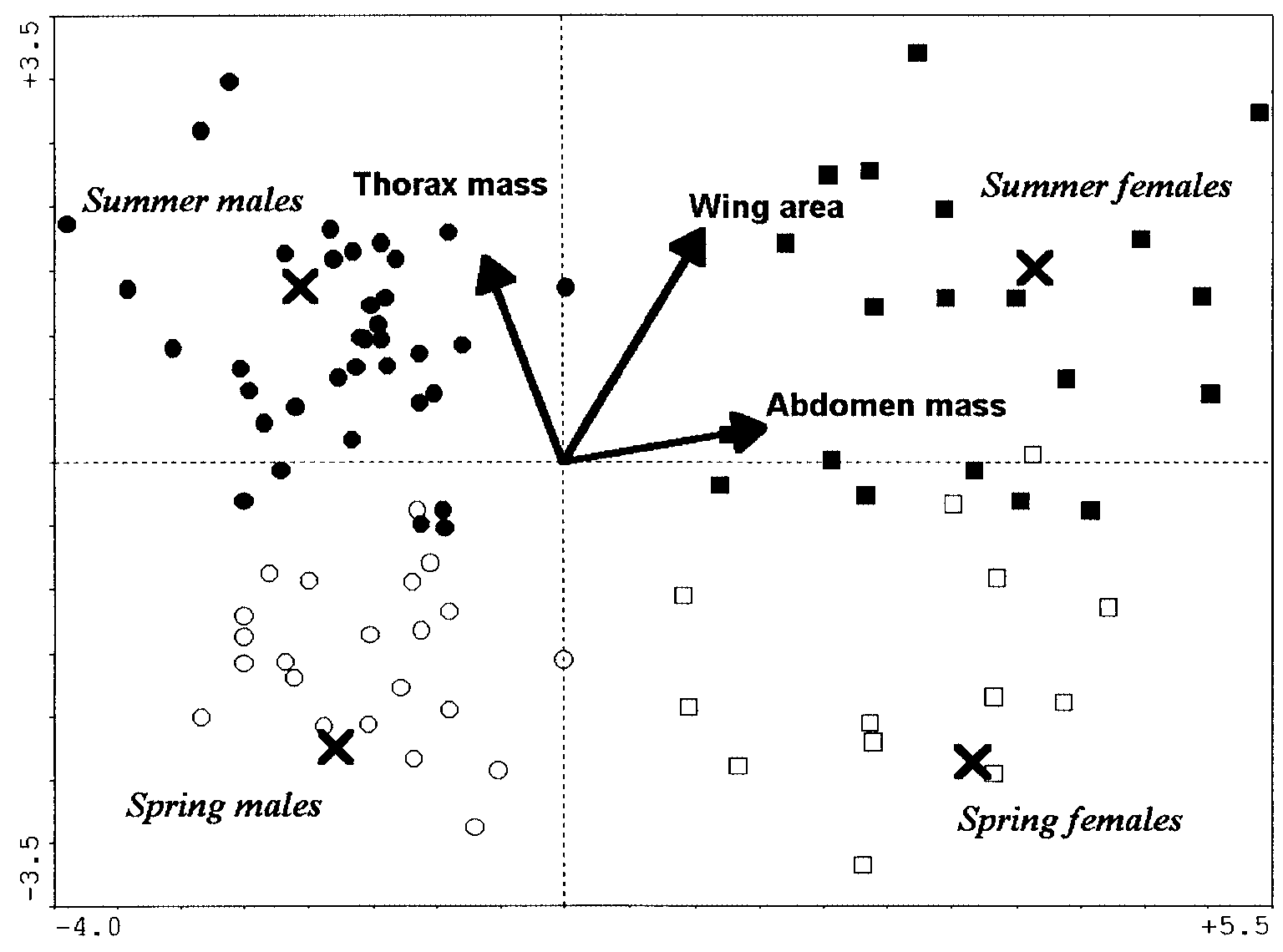 Fig. 2. Canonical variate analysis of biomechanical traits of Araschnia levana. Raw variables, sexes
Fig. 2. Canonical variate analysis of biomechanical traits of Araschnia levana. Raw variables, sexes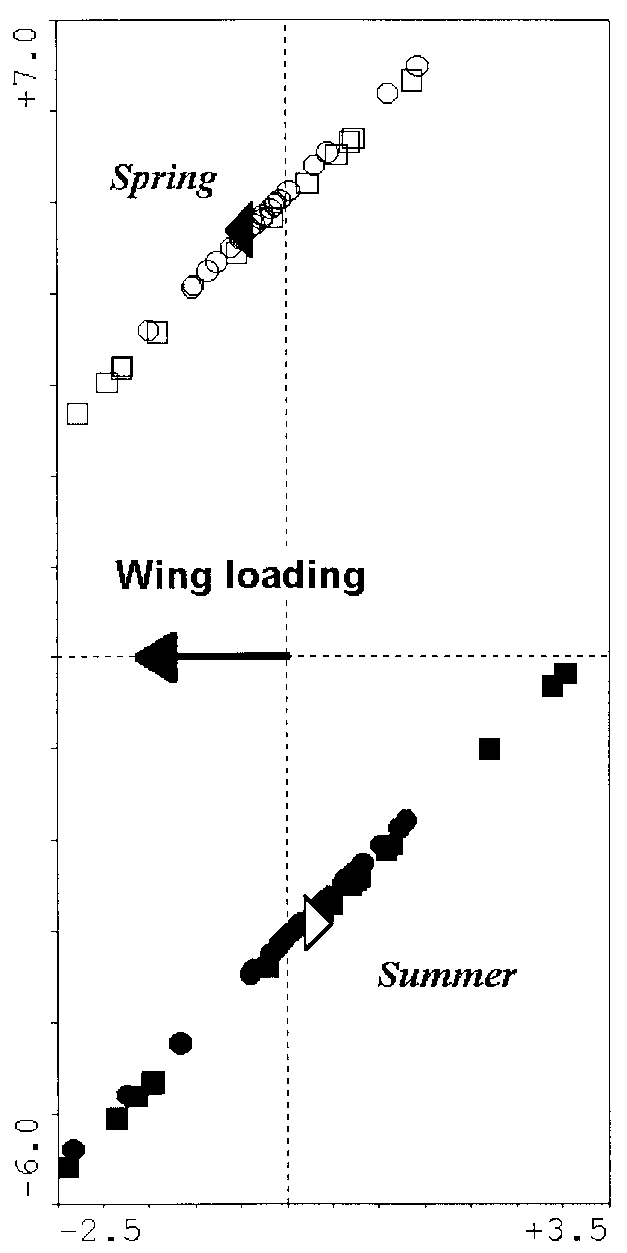 Fig. 3. Canonical variate analysis of biomechanical traits of Araschnia levana. Derived variables,
Fig. 3. Canonical variate analysis of biomechanical traits of Araschnia levana. Derived variables,