Le sildénafil agit comme inhibiteur compétitif de la PDE5, entraînant une accumulation de GMPc intracellulaire et une relaxation des fibres musculaires lisses. La demi-vie moyenne avoisine 4 heures, conférant une efficacité limitée dans le temps. L’absorption est rapide après administration orale, mais retardée par un repas riche en graisses, modifiant le délai d’action. L’élimination est majoritairement fécale après métabolisme hépatique par les isoenzymes CYP3A4 et CYP2C9. Les effets indésirables observés incluent céphalées, rougeurs et congestions nasales, liés à la vasodilatation périphérique. Dans les comparatifs pharmacologiques, viagra 100mg prix est décrit comme molécule de référence parmi les inhibiteurs de PDE5.
Site.trancess.com.my
Effects of Astaxanthin on Lipopolysaccharide-Induced Inflammation In Vitro and In Vivo Kazuhiro Ohgami,1 Kenji Shiratori,1 Satoshi Kotake,1 Tomomi Nishida,2Nobuhisa Mizuki,2 Kazunaga Yazawa,3 and Shigeaki Ohno1
PURPOSE. Astaxanthin (AST) is a carotenoid that is found in
Endotoxin-induced uveitis (EIU) is an animal model of acute
marine animals and vegetables. Several previous studies have
anterior segment intraocular inflammation that is induced
demonstrated that AST exhibits a wide variety of biological
by an injection of lipopolysaccharide (LPS) or lipoteichoic
activities including antioxidant, antitumor, and anti-Helicobac-
acid.1–5 In this model, LPS may directly activate the vascular
ter pylori effects. In this study, attention was focused on the
endothelium, macrophages, and other cells. Cellular infiltration
antioxidant effect of AST. The object of the present study was
and protein extravasation in the anterior part of the eye
to investigate the efficacy of AST in endotoxin-induced uveitis
reaches a maximum at 20 to 24 hours after LPS treatment.6 In
(EIU) in rats. In addition, the effect of AST on endotoxin-
the vitreous and retina, cellular infiltration reaches a maximumat 48 hours after LPS treatment.7 Exposure to outer bacterial
induced nitric oxide (NO), prostaglandin E2 (PGE2), and tumor
toxins such as LPS stimulates cellular inflammatory responses
necrosis factor (TNF)-␣ production in a mouse macrophage
and releases factors, such as nitric oxide (NO),8,9 prostaglandin
cell line (RAW 264.7) was studied in vitro.
E2 (PGE2),10 –12 cytokines including tumor necrosis factor
METHODS. EIU was induced in male Lewis rats by a footpad
(TNF)-␣,13 and eicosanoid mediators, that promote inflamma-
injection of lipopolysaccharide (LPS). AST or prednisolone was
tory responses. In particular, increased plasma TNF-␣ levels
administered intravenously at 30 minutes before, at the same
during endotoxemia and Gram-negative sepsis contributes to
time as, or at 30 minutes after LPS treatment. The number of
lethality as suggested by the protective effects afforded by
infiltrating cells and protein concentration in the aqueous hu-
mor collected at 24 hours after LPS treatment was determined.
Three types of nitric oxide synthase (NOS) isoforms have
RAW 264.7 cells were pretreated with various concentrations
been identified in cells. Endothelium NOS and neural NOS are
of AST for 24 hours and subsequently stimulated with 10
both constitutive NOS. The NO produced by constitutive NOS
g/mL of LPS for 24 hours. The levels of PGE2, TNF-␣, and NO
acts to maintain normal vasoactivity in an active state of vaso-
production were determined in vivo and in vitro.
dilation through a Caϩ2-dependent pathway and acts as aneurotransmitter in neuron signal transmission. NOS in macro-
RESULTS. AST suppressed the development of EIU in a dose-
phages and hepatocytes is inducible (i)NOS and its activation is
dependent fashion. The anti-inflammatory effect of 100 mg/kg
Caϩ2 independent. After exposure to endogenous and exoge-
AST was as strong as that of 10 mg/kg prednisolone. AST also
nous stimulators, iNOS is induced quantitatively in various
decreased production of NO, activity of inducible nitric oxide
cells, such as macrophages, smooth muscle cells, and hepato-
synthase (NOS), and production of PGE2 and TNF-␣ in
cytes to trigger several disadvantageous cellular responses and
RAW264.7 cells in vitro in a dose-dependent manner.
cause inflammation.15 Therefore, NO production induced by
CONCLUSIONS. This study suggests that AST has a dose-depen-
iNOS may reflect the degree of inflammation. Thus, we can
dent ocular anti-inflammatory effect, by the suppression of NO,
evaluate the effect of an anti-inflammatory drug by measuring
PGE2, and TNF-␣ production, through directly blocking NOS
NO levels. N-nitro-L-arginine methyl ester (L-NAME) showedeffective inhibitory activity in LPS-induced NO production by
directly blocking the NOS enzyme activities.16 –18
Carotenoids are a family of more than 700 natural lipid-
soluble pigments that are only produced by phytoplankton,algae, plants, and a limited number of fungi and bacteria.
From the 1Department of Ophthalmology and Visual Sciences,
Astaxanthin (AST) is one of the most common carotenoids and
Hokkaido University Graduate School of Medicine, Sapporo, Japan; the
is found in the red pigment of crustacean shells (crabs,
2Department of Ophthalmology, Yokohama City University School of
shrimps, for example), salmon, and the asteroidean.19 The
Medicine, Yokohama, Japan; and the 3Laboratory of Nutraceuticals and
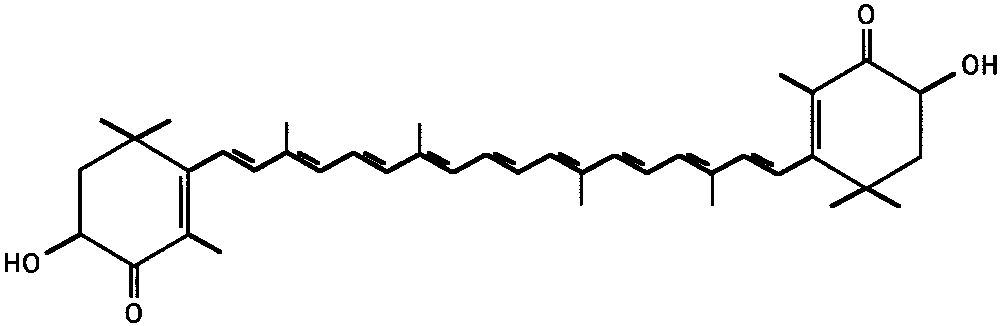
chemical structure of AST is shown in Figure 1. The AST of the
Functional Foods Science, Graduate School of Fisheries Science, Tokyo
xanthophylls group possesses no provitamin A activity in con-
University of Fisheries, Tokyo, Japan.
trast to ␣-carotene. The AST content of salmon is 1.7 to 2.6
Supported in part by a grant for Researches on Sensory and
mg/100 g. Several previous studies have demonstrated that AST
Communicative Disorders, Ministry of Health, Labour and Welfare,
exhibits a wide variety of biological activities, including anti-
Japan, and in part by grants-in-aid for Scientific Research, Ministry of
oxidant,20 antitumor,21 and anti-Helicobacter pylori effects.22
Education, Culture and Science, Japan.
Submitted for publication August 14, 2002; revised December 16,
The antioxidant activities of carotenoids are related to the
stability of formed free radicals after they react with active free
Disclosure: K. Ohgami, None; K. Shiratori, None; S. Kotake,
radicals. As a result, AST could suppress the production of NO.
None; T. Nishida, None; N. Mizuki, None; K. Yazawa, None; S.
Several in vitro and in vivo studies have indicated that L-NAME
Ohno, None
may also function as an anti-inflammatory mediator.16,18 There
The publication costs of this article were defrayed in part by page
has been no report on the effects of AST on LPS-induced
charge payment. This article must therefore be marked “advertise-ment” in accordance with 18 U.S.C. §1734 solely to indicate this fact.
In the present study, we investigated the influence of AST
Corresponding author: Kazuhiro Ohgami, Department of Ophthal-
on LPS-induced uveitis in rats. In addition, we also investigated
mology and Visual Sciences, Hokkaido University School of Medicine,N17 W5, Kita-ku, Sapporo, 060-8638 Japan;
the NO production in RAW 264.7 cells treated with AST in
vitro to clarify the anti-inflammatory effect. Furthermore, in
Investigative Ophthalmology & Visual Science, June 2003, Vol. 44, No. 6
Copyright Association for Research in Vision and Ophthalmology
Anti-inflammatory Effect of Astaxanthin
ous humor samples was measured with a bicinchoninic acid (BCA)protein assay kit (Pierce, Rockford, IL). The aqueous humor sampleswere stored in ice water until testing, and cell counts, and total proteinconcentrations were measured on the day of sample collection. Determination of NO Levels in Aqueous Humor
The total level of nitrate plus nitrite in the aqueous humor was mea-sured by using a total nitrite colorimetric assay kit (Oxis International,Portland, OR), according to the manufacturer’s instruction. The aque-
FIGURE 1.
ous humor from both eyes of a rat was diluted up to 50 L and usedfor one assay. The NO assay was repeated once or twice.
vivo, the anti-inflammatory potency of AST was compared withthat of prednisolone. Also, in vitro, the inhibitory effect of AST
Levels of TNF-␣ and PGE2 in Aqueous Humor
on NO production was compared with that of L-NANE, aknown inhibitor of NO production.
The levels of TNF-␣ and PGE2 in the aqueous humor obtained from ratswith EIU were assessed with a commercially available ELISA kit (R&DSystems, Minneapolis, MN), according to the manufacturer’s instruc-
MATERIALS AND METHODS
tions. The ELISA assay was performed in duplicate. The data representthe mean of eight determinations Ϯ SD. Animals Groups and EIU Induction
Eight-week-old male Lewis rats, weighing 180 to 220 g, were used. EIU
Cell Culture and LPS Stimulation
was induced by injection into one footpad of 200 g of LPS from
RAW 264.7, a mouse macrophage cell line, was obtained from the
Salmonella typhimurium (Sigma, St. Louis, MO) that had been diluted
American Type Culture Collection (Manassas, VA). Cells were cultured
in RPMI-1640 medium supplemented with 2 mM glutamine, antibiotics
The rats were injected intravenously with 1, 10, or 100 mg/kg AST
(100 U/mL each of penicillin and streptomycin), and 10% heat-inacti-
(Sigma) or 10 mg/kg prednisolone (Sigma) in 1 mL/kg 60% polyethyl-
vated fetal bovine serum (Gibco-BRL, Grand Island, NY) and main-
ene glycol (Wako, Osaka, Japan). Each compound was administered a
tained at 37°C in a humidified incubator containing 5% CO . RAW
three time points: simultaneously and 30 minutes before and after the
264.7 cells were seeded onto a 24-well plate (5 ϫ 104 cells/well) for
LPS injection. For the LPS group, 60% polyethylene glycol was admin-
experiments. The cells were pretreated with 2.5, 5, 12.5, and 25 M
istered intravenously on the same schedule as the AST group.
AST for 24 hours and subsequently stimulated with 10 g/mL of LPS
Animals were handled and cared for according to the ARVO State-
from S. typhimurium for 24 hours, unless otherwise stated.
ment for the Use of Animals in Ophthalmic and Vision Research.
AST was dissolved in 0.01% dimethyl sulfoxide (DMSO). For the
Number of Infiltrating Cells and Protein
control group, RAW cells were cultured with 0.01% DMSO alone. Theeffects of AST were compared with those of
Concentration in Aqueous Humor
Suzuma reported that cellular infiltration in the aqueous humorreached a maximum level at 24 hours after LPS treatment in this
Determination of Nitrite Concentration
model.1 The number of cells infiltrating the aqueous humor and the
in Medium
aqueous humor protein concentration were used as indicators of the
NO was measured as its end product, nitrite, by using Griess reagent
(Sigma), as described elsewhere.23 The culture supernatant (100 L)
At 24 hours after LPS injection, rats were killed and the aqueous
was mixed with 100 L of Griess reagent for 10 minutes, and the
humor was collected immediately. Briefly, the aqueous humor was
absorbance at 550 nm was measured in a microplate reader. The
collected from both eyes by an anterior chamber puncture (15–20
concentration of nitrite in the samples was determined with reference
L/rat) with a 30-gauge needle under a surgical microscope. For cell
to a sodium nitrite standard curve. The data represent the mean of
counting, the aqueous humor sample was suspended in an equal
¨rk stain solution, and the cells were counted, using a
hemocytometer under a light microscope. The number of cells per
Determination of iNOS Enzyme Activity
field (an equivalent of 0.1 mL) was manually counted, and the numberof cells per microliter was obtained by averaging the results of four
A NO synthase assay kit (Calbiochem-Novabiochem, San Diego, CA)
fields from each sample. The total protein concentration in the aque-
was used to determine iNOS enzyme activity. AST- or L-NAME–pre-
FIGURE 2.
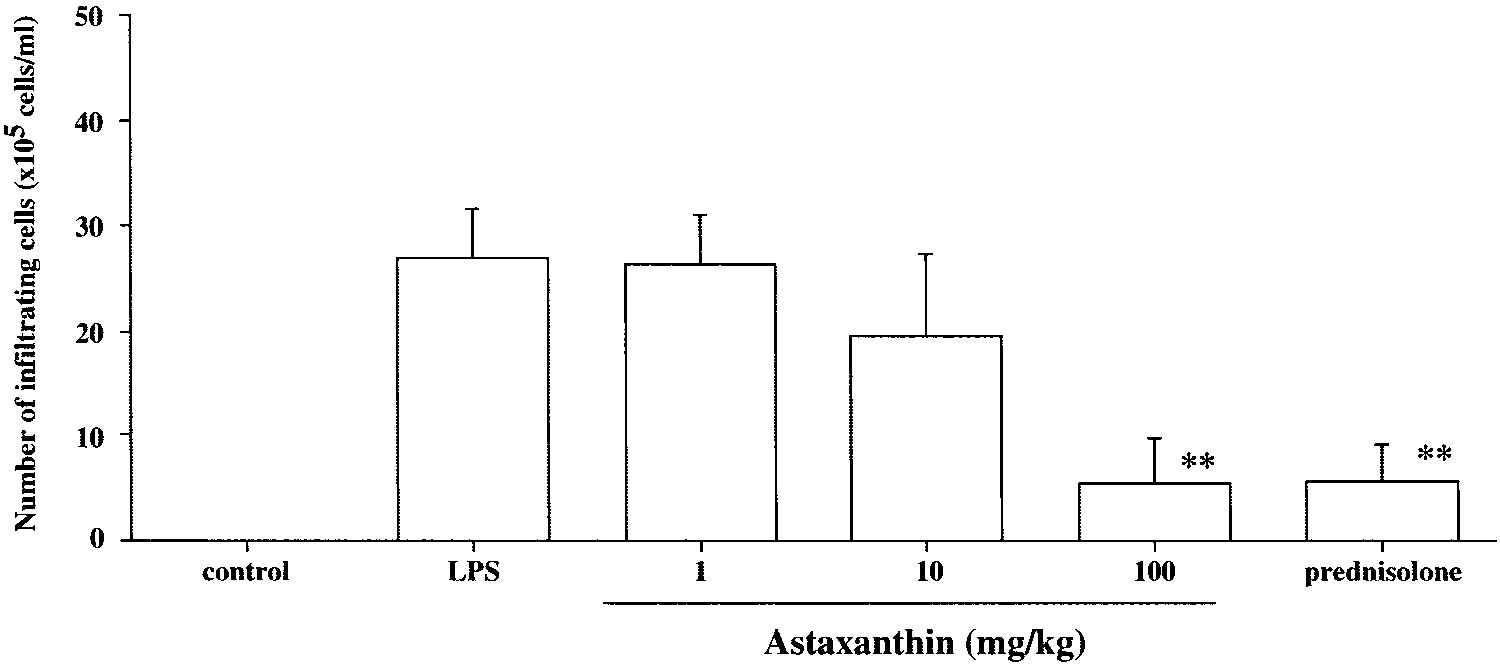
cellular infiltration in the aqueous hu-mor. The aqueous humor was col-lected 24 hours after LPS treatment. Cell numbers are expressed as themean Ϯ SD (n ϭ 8). No infiltratingcells were detected in the aqueoushumor from rats without LPS (con-trol group). The dose of prednisolonewas 10 mg/kg. **P Ͻ 0.01, comparedwith the control group. Ohgami et al. FIGURE 3.
protein concentration in the aque-ous humor. The aqueous humorwas collected 24 hours after LPStreatment. Each value representsthe mean Ϯ SD (n ϭ 8). The dose ofprednisolone was 10 mg/kg. *P Ͻ0.05 and **P Ͻ 0.01, comparedwith the LPS group.
treated cells were incubated with LPS (10 g/mL) for 24 hours. The
cells were washed three times with PBS, scraped into cold PBS, andcentrifuged at 500g for 10 minutes at 4°C. The cell pellet was resus-
Number of Inflammatory Cells
pended in 0.4 mL of hypotonic buffer that contained 10 mM HEPES, 10
in Aqueous Humor
mM KCl, 1 mM dithiothreitol, 1 mM phenylmethylsulfonyl fluoride,
In the LPS group, the number of inflammatory cells that infil-
and 0.2 mM EDTA (pH 7.4). The total protein concentrations in
trated the aqueous humor 24 hours after LPS treatment was
solution samples were measured using a BCA protein assay kit (Pierce).
26.9 Ϯ 4.7 ϫ 105 cells/mL (mean Ϯ SD, n ϭ 7). The grouptreated with 100 mg/kg of AST showed a significantly reduced
Levels of TNF-␣ and PGE2
number of inflammatory cells (5.4 Ϯ 4.3 ϫ 105 cells/mL)compared with the control group (P Ͻ 0.01, Fig. 2). The effect
The levels of TNF-␣ and PGE2 in the medium were measured by ELISA
of 100 mg/kg AST on the number of cells in the aqueous humor
(R&D Systems) according to the manufacturer’s instruction. The ELISA
was almost the same as that for 10 mg/kg prednisolone (5.7 Ϯ
4.3 ϫ 105 cells/mL, Fig. 2). Treatment with 10 mg/kg of ASTshowed a mild reduction in number of cells (19.6 Ϯ 7.7 ϫ 105
Cell Viability
cells/mL), and there was no significant difference from the LPSgroup. No infiltrating cells were detected in the aqueous hu-
For determination of cell viability, 50 mg/mL of methylthiazol-2-yl-2,5-
mor from rats without LPS (control group).
diphenyl tetrazolium bromide (Sigma) was added to 1 mL of cellsuspension (5.3 ϫ 104 cells/mL in 24-well plates) for 24 hours, and the
Aqueous Humor Protein Concentration
MTT formazan formed was dissolved in acidic-2-propanol. Opticaldensity was measured with a plate reader used at 590 nm. The optical
The protein concentration in the aqueous humor of rats with-
density of the Formosan formed by untreated cells was taken as 100%.
out LPS (control group) was 2.0 Ϯ 0.5 mg/mL and in that ofrats with LPS was 20.1 Ϯ 2.8 mg/mL. The protein concentra-
Statistical Analysis
tions in the groups treated with 10 mg/kg and 100 mg/kg ASTwere significantly lower than that in the LPS group (10 mg/kg:
The values were expressed as mean Ϯ SD. A Student’s unpaired t-test
15.3 Ϯ 2.6 mg/mL, P Ͻ 0.05; 100 mg/kg: 6.2 Ϯ 1.7 mg/mL, P Ͻ
was used to assess the statistical significance of differences. P Ͻ 0.05
0.01, Fig. 3). The reduction of protein concentration in the AST
100-mg/kg group was almost the same as that in the pred-
FIGURE 4.
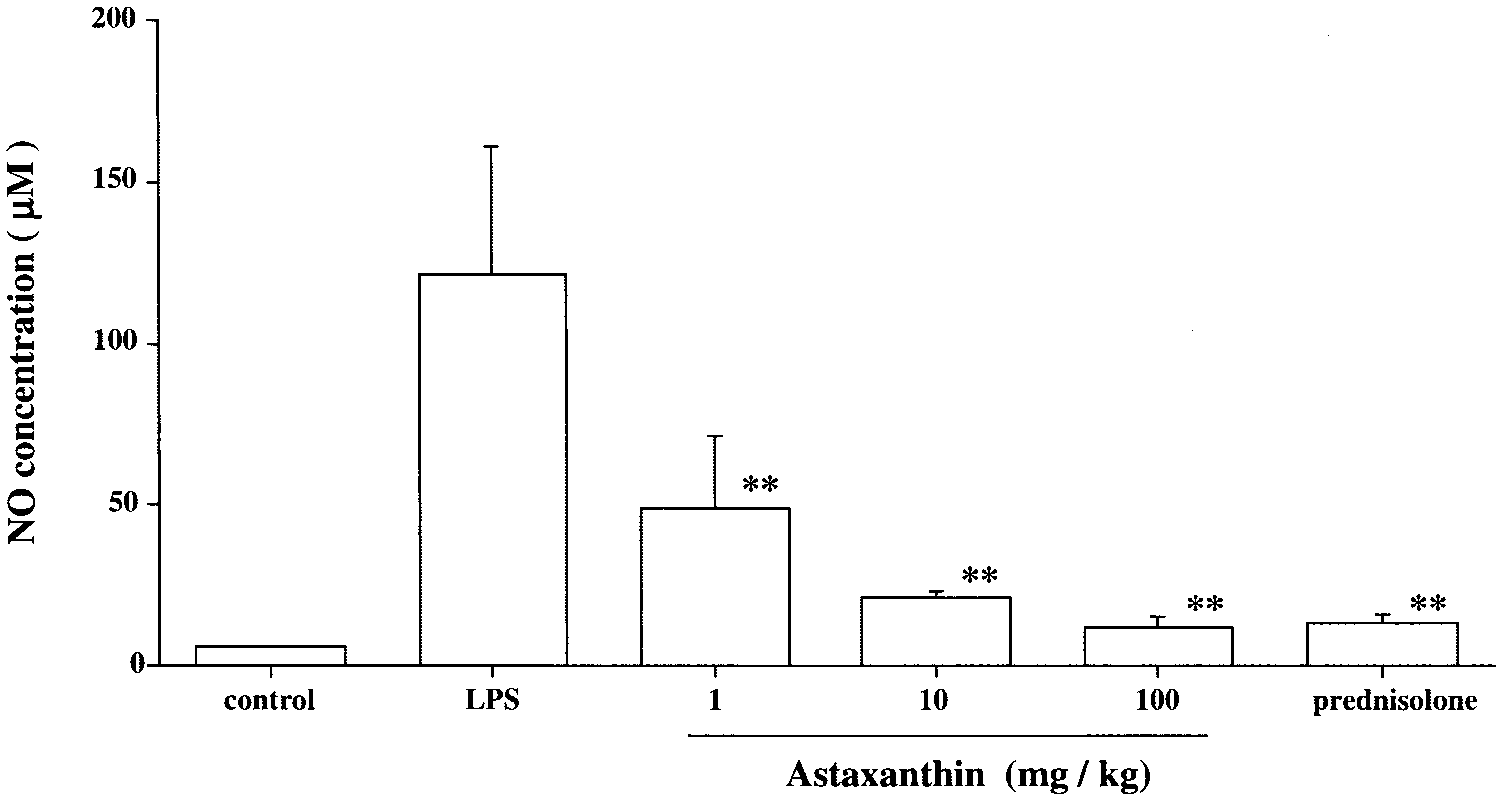
NO levels in the aqueous humor. Theaqueous humor was collected 24hours after LPS treatment. Each valuerepresents mean Ϯ SD (n ϭ 8). Thedose of prednisolone was 10 mg/kg. **P Ͻ 0.01, compared with the LPSgroup. Anti-inflammatory Effect of Astaxanthin FIGURE 5.
TNF-␣ concentrations in the aqueoushumor. The aqueous humor was col-lected 24 hours after LPS treatment. Each value represents the mean Ϯ SD(n ϭ 8). The dose of prednisolonewas 10 mg/kg. **P Ͻ 0.01, comparedwith the LPS group.
nisolone group (6.5 Ϯ 2.3 mg/mL, Fig. 3). Treatment with 1
the AST group showed a tendency to decrease in a dose-
mg/kg of AST produced only a mild reduction in protein
dependent fashion. Treatment with AST significantly reduced
concentration (17.7 Ϯ 3.2 mg/mL), and there was no signifi-
the TNF-␣ concentration compared with that of the LPS group
cant difference from level in the LPS group.
(1 mg/kg: 287.6 Ϯ 42.8 pg/mL, P Ͻ 0.01; 10 mg/kg: 241.0 Ϯ27.5 pg/mL, P Ͻ 0.01; 100 mg/kg: 223.1 Ϯ 24.3 pg/mL, P Ͻ
Levels of NO in EIU
0.01, Fig. 5). The reduction in TNF-␣ concentration in the AST
The NO production in the LPS group was 122.0 Ϯ 39.1 M (n
100-mg/kg group was almost the same as that in the pred-
ϭ 8). The NO production in the groups treated with AST
nisolone group (233.0 Ϯ 2.3 pg/mL, Fig. 5). However, TNF-␣
differed significantly from that in the LPS control group, in a
was not detected in the aqueous humor of the control rats.
dose-dependent manner. (1 mg/kg: 49.1 Ϯ 23.2 M, P Ͻ 0.01;10 mg/kg: 21.1 Ϯ 2.3 mg/mL, P Ͻ 0.01; 100 mg/kg: 12.1 Ϯ 2.9
PGE2 Concentration in EIU
M, P Ͻ 0.01; Fig. 4). The reduction in NO production in the
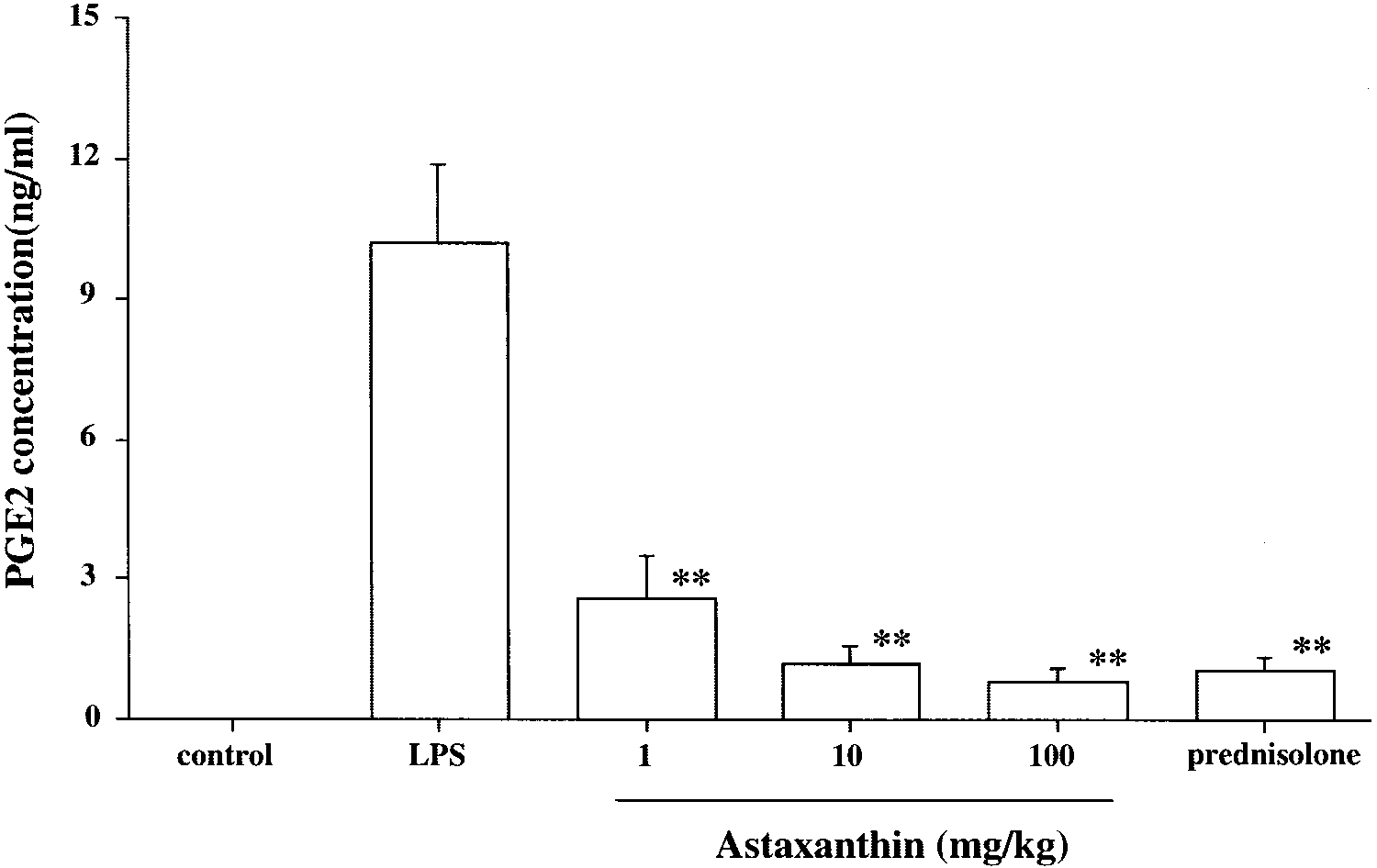
The PGE2 concentration in the LPS control group was 10.2 Ϯ
AST 100-mg/kg group was almost the same as that in the
1.7 ng/mL. Treatment with AST significantly reduced PGE2
prednisolone group (13.3 Ϯ 3.2 M, Fig. 4). The NO concen-
concentration, compared with that in the LPS group (1 mg/kg:
tration in the aqueous humor of rats without LPS (control
2.6 Ϯ 0.9 ng/mL, P Ͻ 0.01; 10 mg/kg: 1.2 Ϯ 0.4 ng/mL, P Ͻ
0.01; 100 mg/kg: 0.8 Ϯ 0.3 ng/mL, P Ͻ 0.01; Fig. 6). Thereduction in PGE2 concentration in the AST 100-mg/kg group
Levels of TNF-␣ in Aqueous Humor
was almost the same as that in the prednisolone group (1.0 Ϯ
In the LPS group, the TNF-␣ concentration in the aqueous
0.3 ng/mL, Fig. 6). PGE2 was not detected in the aqueous
humor was 792.3 Ϯ 140.1 pg/mL. The TNF-␣ concentration in
FIGURE 6.
PGE2 concentrations in the aqueoushumor. The aqueous humor was col-lected 24 hours after LPS treatment. Each value represents the mean Ϯ SD(n ϭ 8). The dose of prednisolonewas 10 mg/kg. **P Ͻ 0.01, comparedwith the LPS group. Ohgami et al. FIGURE 7.
NO production in LPS- induced RAW264.7 cells. RAW cells were pre-treated with various concentrationsof astaxanthin and L-NAME for 24hours. Astaxanthin- or L-NAME–pre-treated RAW cells were incubatedwith LPS (10 g/mL) for 24 hours. The nitrite concentration in the cul-ture supernatants was determinedfor NO production. RAW cells in thenormal group were cultured with0.01% DMSO. Data are expressed asthe mean Ϯ SD (n ϭ 8). **P Ͻ 0.01,compared with the LPS group. NO Production in RAW 264.7 Cells
and 25 M: 0.66 Ϯ 0.22 nmol/min per milligram protein, P Ͻ0.01, Fig. 8). The effect of 25 M AST on LPS-induced NO
The content of nitrite without LPS in RAW 264.7 cells (control
production in RAW 264.7 cells was almost the same as that of
group) was 4.2 Ϯ 0.9 M (n ϭ 8). After the treatment with 10
g/mL of LPS for 24 hours (LPS group), nitrite concentration in
the medium increased substantially, by approximately 10-fold
TNF-␣ Concentration
(41.7 Ϯ 4.4 M). Treatment with 2.5, 12.5, or 25 M ASTsignificantly reduced NO production compared with that in
The TNF-␣ concentration induced by LPS was approximately
the LPS group (2.5 M: 28.7 Ϯ 6.5 M, P Ͻ 0.01; 12.5 M:
10 times higher than in the normal group. The TNF-␣ concen-
15.6 Ϯ 1.4 M, P Ͻ 0.01; 25 M: 11.5 Ϯ 2.5 M, P Ͻ 0.01; Fig.
tration in the AST group showed a tendency to decrease in a dose-
7). The effect of 25 M AST on LPS-induced NO production in
dependent manner. Treatment with AST significantly reduced
RAW 264.7 cells was almost the same as that of 25 M L-NAME
TNF-␣ concentration compared with that in the LPS group (5
(10.3 Ϯ 1.6 M, Fig. 7). AST did not decrease cell viability in
M: 893.0 Ϯ 95.0 pg/mL, P Ͻ 0.05; 12.5 M: 501.8 Ϯ 86.7
RAW 264.7 cells when these cells were incubated with 25 M
pg/mL, P Ͻ 0.01; and 25 M: 375.3 Ϯ 35.9 pg/mL, P Ͻ 0.01;
AST alone for 24 hours (data not shown).
Fig. 9). Treatment with L-NAME significantly reduced TNF-␣levels compared with those of the LPS group. However, the
iNOS Enzyme Activity
TNF-␣ levels of the 25-M AST group was significantly lowerthan those of the 25-M
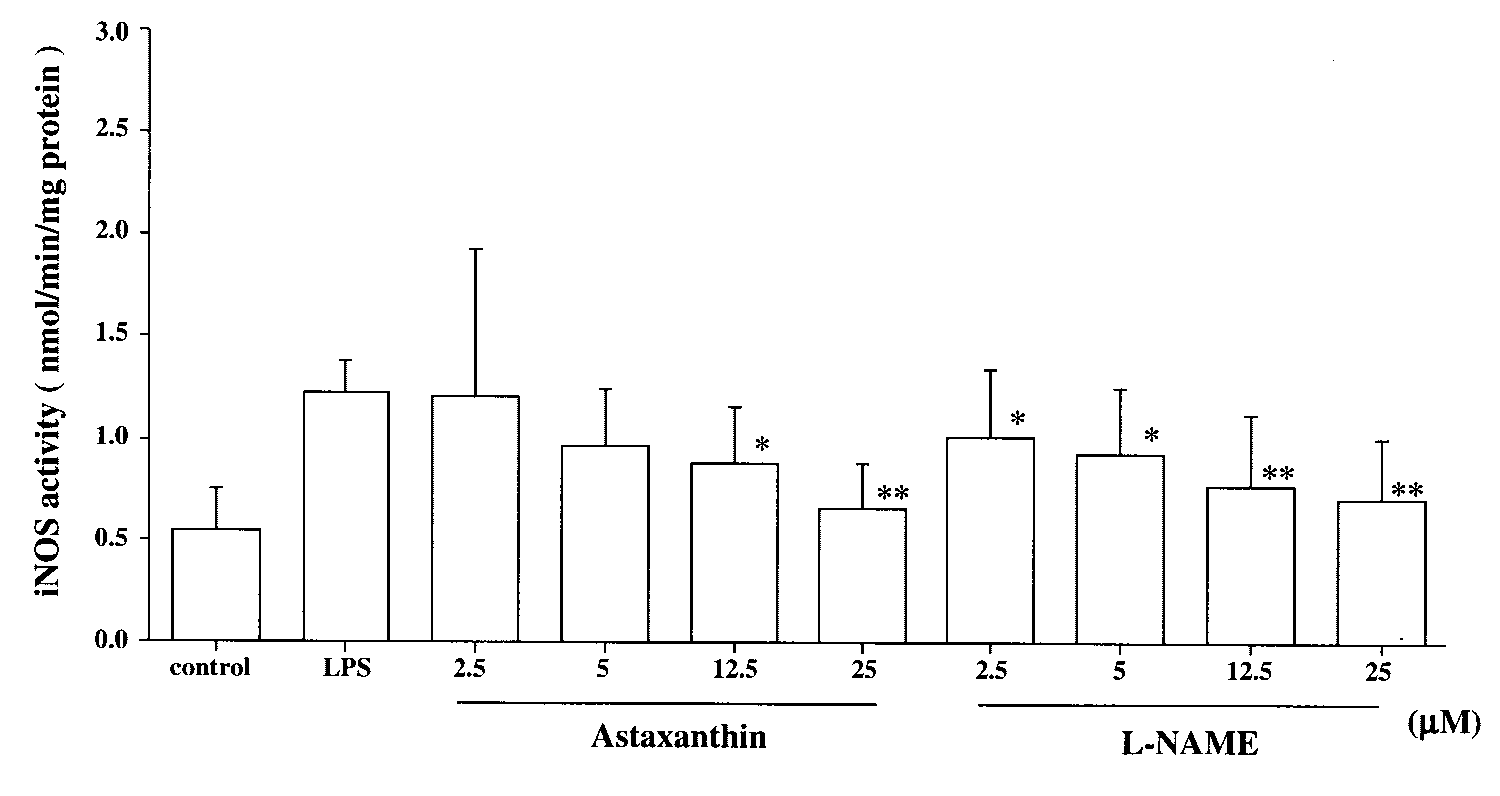
NOS enzyme activity in RAW 264.7 cells without stimulation
was 0.54 Ϯ 0.20 nmol/min per milligram protein (n ϭ 8). After
L-NAME; 787.0 Ϯ 117.0 pg/mL, P Ͻ 0.01, Fig. 9).
LPS stimulation, the NOS enzyme activity was approximately
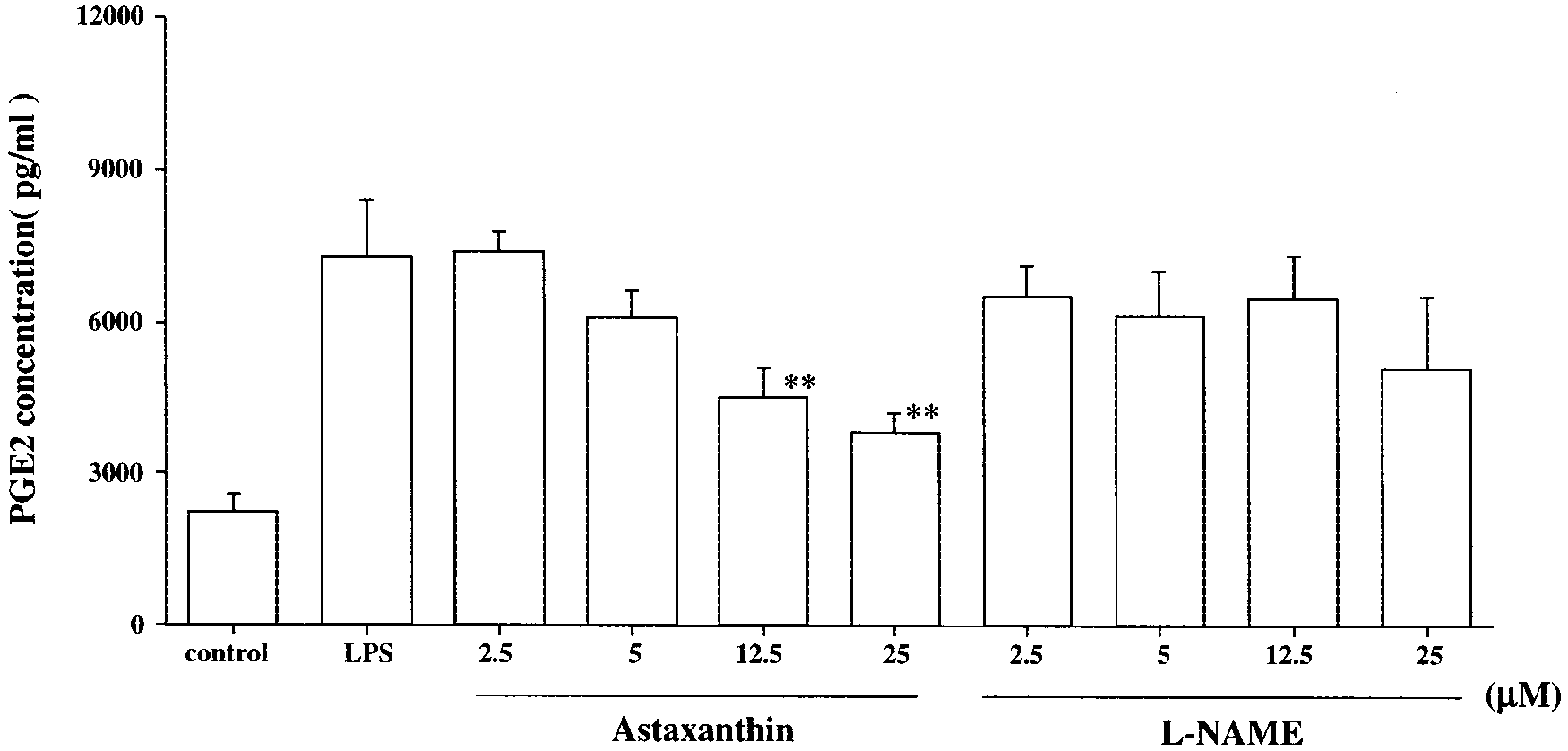
PGE2 Concentration
two times higher than in the normal group (1.23 Ϯ 0.16nmol/min per milligram protein). Enzyme activity showed a
The PGE2 concentration induced by LPS was approximately
tendency to decrease in the AST-treated group in a dose-
four times higher than in the normal group. The PGE2 concen-
dependent manner. Treatment with AST significantly reduced
tration in the AST group showed a tendency to decrease in
iNOS activity compared with that in the LPS group (5 M:
conjunction with decreases in AST concentrations. Treatment
0.97 Ϯ 0.28 nmol/min per milligram protein; P Ͻ 0.05, 12.5
with AST significantly reduced PGE2 concentration compared
M: 0.88 Ϯ 0.28 nmol/min per milligram protein, P Ͻ 0.01;
with that in the LPS group (12.5 M: 4529.4 Ϯ 598.2 pg/mL,
FIGURE 8.
NOS activity in LPS-induced RAW264.7 cells. RAW cells were pre-treated with various concentrationsof astaxanthin and L-NAME for 24hours. Astaxanthin- or L-NAME–pre-treated RAW cells were incubatedwith LPS (10 g/mL) for 24 hours. RAW cells in the normal group werecultured with 0.01% DMSO. Data areexpressed as the mean Ϯ SD (n ϭ 8). *P Ͻ 0.05 and **P Ͻ 0.01, comparedwith the LPS group. Anti-inflammatory Effect of Astaxanthin FIGURE 9.
Effect of astaxanthin on TNF-␣ levels in LPS-infected RAW 264.7 cells. RAW cells were pretreated with various concentrations of
astaxanthin and L-NAME for 24 hours. Astaxanthin- or L-NAME-pretreated RAW cells were incubated with LPS (10 g/mL) for 24 hours. RAW cellsof normal group were cultured with 0.01% DMSO. Data are expressed as the mean Ϯ SD (n ϭ 8). *P Ͻ 0.05 and **P Ͻ 0.01, compared with theLPS group. P Ͻ 0.01; and 25 M: 3797.7 Ϯ 401.7 pg/mL, P Ͻ 0.01; Fig.
against damage from singlet oxygen in vitro23— 80 times stron-
10). Treatment with L-NANE showed mild PGE2 concentration
ger than ␣-tocopherol and twice as strong as -carotene.22,23 It
reduction and there were no significant differences from the
is understandable why AST has a strong singlet oxygen-quench-
ing capability, considering its molecular structure. The reactiv-ity to other molecular oxygen decreases, because singlet-oxy-
DISCUSSION
gen–associated, carbon-centered radicals of AST can form morestable resonance structures by the attachment of the carbonyl
The results of this study indicate that AST suppresses the
group and the hydroxyl group to the -ionone ring of AST.23
development of EIU in a dose-dependent fashion. In particular,
AST can remove the chain carrying lipid peroxyl radicals in the
the ocular anti-inflammatory effect of 100 mg/kg of AST was as
liposomal suspension more efficiently than -carotene but less
strong as that of a 10-mg/kg dose of prednisolone.
efficiently than ␣-tocopherol, because the hydrogen bonds by
Carotenoids are known to take part in protecting marine
the carbonyl group in the -ionone ring of AST and hydropho-
animals against damage from free radicals and singlet oxygen
bic association by the polyene chain allows AST to fit in the
reactive species.22 AST has a strong quenching capability
membrane phospholipid structure well.24 –27
FIGURE 10.
Effect of astaxanthin on PGE2 concentrations in LPS-infected RAW 264.7 cells. RAW cells were pretreated with various concentrations
of astaxanthin and L-NAME for 24 hours. Astaxanthin- or L-NAME–pretreated RAW 264.7 cells were incubated with LPS (10 g/mL) for 24 hours. RAW 264.7 cells of the normal group were cultured with 0.01% DMSO. Data are expressed as the mean Ϯ SD (n ϭ 8). **P Ͻ 0.01, compared withthe LPS group. Ohgami et al.
To elucidate the anti-inflammatory mechanism of AST, we
7. Miyamoto K, Ogura Y, Hamada M, Nishiwaki H, Hiroshiba N,
focused our attention on the antioxidant effect of AST and
Honda Y. In vivo quantification of leukocyte behavior in the retina
measured the concentration of NO in the aqueous humor in
during endotoxin-induced uveitis. Invest Ophthalmol Vis Sci.
vivo. AST suppressed the NO production in the aqueous hu-
mor in a dose-dependent manner. Also, we investigated the
8. Chen YC, Shen SC, Lee WR, Hou WC, Yang LL, Lee TJ. Inhibition
effect of AST on LPS-induced NO production and PGE2 and
of nitric oxide synthase inhibitors and lipopolysaccharide inducedinducible NOS and cyclooxygenase-2 gene expressions by rutin,
TNF-␣ levels in the RAW 264.7 macrophage cell, with L-NAME
quercetin, and quercetin pentaacetate in RAW 264.7 macro-
as a positive control. AST decreased NO production and iNOS
phages. J Cell Biochem. 2001;82:537–548.
enzyme activity in a dose-dependent manner, thus agreeing
9. Boujedaini N, Liu J, Thuillez C, Cazin L, Mensah-Nyagan AG. In vivo
with the results of the in vivo experiment. These results dem-
regulation of vasomotoricity by nitric oxide and prostanoids dur-
onstrate that AST suppresses NO production by directly inhib-
ing gestation. Eur J Pharmacol. 2001;427:143–149.
iting the NOS enzyme activity similar to the NOS inhibitor
10. Bellot JL, Palmero M, Garcı´a-Cabanes C, Espı´ R, Hariton C, Orts A.
L-NAME. Large amounts of NO production induced by bacterial
Additive effect of nitric oxide and prostaglandin-E2 synthesis in-
lipopolysaccharide or cytokines play an important role in en-
hibitors in endotoxin-induced uveitis in the rabbit. Inflamm Res.
dotoxemia and inflammatory conditions.10 Therefore, we pro-
pose that AST, which inhibits NO production through inhibit-
11. Murakami A, Nakamura Y, Tanaka T, et al. Suppression by citrus
ing iNOS enzyme activity, has beneficial therapeutic effects in
auraptene of phorbol ester- and endotoxin-induced inflammatory
the treatment of inflammation. Our results indicate that the com-
responses: role of attenuation of leukocyte activation. Carcinogen-
pound, even at the concentration of 25 ⌴, did not change cell
viability. Therefore, inhibition of LPS-induced NO production by
12. Hoekzema R, Verhagen C, van Haren M, Kijlstra A. Endotoxin-
AST was not the result of its cytotoxicity on the cells.
induced uveitis in the rat: the significance of intraocular interleu-kin-6. Invest Ophthalmol Vis Sci. 1992;33:532–539.
TNF-␣ is a pleiotropic cytokine produced principally by
13. Tracey KJ, Cerami A. Tumor necrosis factor: a pleiotropic cytokine
activated macrophages and monocytes and also has an major
and therapeutic target. Annu Rev Med. 1994;45:491–503.
role in the nonspecific resistance against various infectious
14. Tracey KJ, Fong Y, Hesse DG, et al. Anti-cachectin/TNF monoclo-
agents.28,29 The results of the present study indicate that AST
nal antibodies prevent septic shock during lethal bacteraemia.
decreased TNF-␣ concentration in a dose-dependent manner,
both in vivo and in vitro. The results of TNF-␣ inhibition by AST
15. Wadsworth TL, Koop DR. Effects of Ginkgo biloba extract (EGb
correspond to the results of inhibited NOS activity and de-
761) and quercetin on lipopolysaccharide-induced release of nitric
creased NO production, with the application of AST.
oxide. Chem Biol Interact. 2001;137:43–58.
The mechanism of the NO-induced suppression of TNF
16. Rahman MM, Bhoola KD, Elson CJ, Lemon M, Dieppe PA. Identi-
synthesis is not known. A potential link is PGE2. It has been
fication and functional importance of plasma kallikrein in the
reported recently that NO activates cyclooxygenase enzymes
synovial fluids of patients with rheumatoid, psoriatic, and osteo-
and thereby leads to a marked increase in PGE2 production.30,31
arthritis. Ann Rheum Dis. 1995;54:345–350.
A suppressive effect of PGE2 on TNF synthesis through elevated
17. Kluft C. Determination of prekallikrein in human plasma: optimal condi-
cAMP levels has been convincingly demonstrated.28,31–33 In the
tions for activating prekallikrein. J Lab Clin Med. 1978;91:83–95.
present study, AST suppressed the levels of LPS-induced PGE2 and
18. Pastor C, Teisseire B, Vicaut E, Payen D. Effects of L-arginine and
TNF-␣ in a dose-dependent manner in vivo and in vitro. Our
L-nitro-arginine treatment on blood pressure and cardiac output in
results support the argument for a regulatory role of NO on TNF
a rabbit endotoxin shock model. Crit Care Med. 1994;22:465– 469.
production in the pathophysiology of EIU.
19. Miki W, Yamaguchi K, Konosu S. Comparison of carotenoids in the
In summary, in the current study AST had a dose-dependent
ovaries of marine fish and shellfish. Comp Biochem Physiol B. 1982;71:7–11.
anti-inflammatory effect on EIU. In particular, the ocular anti-
20. Esterbauer H, Jurgens G, Quehenberger O, Koller E. Autoxidation
inflammatory effect of 100 mg/kg of AST was as strong as that
of human low density lipoprotein: loss of polyunsaturated fatty
of 10 mg/kg prednisolone. A possible mechanism for the ocu-
acids and vitamin E and generation of aldehydes. J Lipid Res.
lar anti-inflammatory effect of AST is the suppression of pro-
duction of NO, PGE2, and TNF-␣ by directly blocking NOS
21. Tanaka T, Makita H, Ohnishi M, Mori H, Satoh K, Hara A. Chemo-
enzyme activity. These results suggest that AST may be a
prevention of rat oral carcinogenesis by naturally occurring xan-
promising agent for the treatment of ocular inflammation.
thophylls, astaxanthin and canthaxanthin. Cancer Res. 1995;55:4059 – 4064.
22. Wang X, Willen R, Wadstrom T. Astaxanthin-rich algal meal and
References
vitamin C inhibit Helicobacter pylori infection in BALB/cA mice.
1. Suzuma I, Mandai M, Suzuma K, Ishida K, Tojo SJ, Honda Y. Antimicrob Agents Chemother. 2000;44:2452–2457.
Contribution of E-selectin to cellular infiltration during endotoxin-
23. Sewer MB, Barclay TB, Morgan ET. Down-regulation of cyto-
induced uveitis. Invest Ophthalmol Vis Sci. 1998;39:1620 –1630.
chrome P450 mRNAs and proteins in mice lacking a functional
2. Hikita N, Chan CC, Whitcup SM, Nussenblatt RB, Mochizuki M.
NOS2 gene. Mol Pharmacol. 1998;54:273–279.
Effects of topical FK506 on endotoxin-induced uveitis (EIU) in the
24. Palozza P, Krinsky NI. Antioxidant effects of carotenoids in vivo
Lewis rat. Curr Eye Res. 1995;14:209 –214.
and in vitro: an overview. Methods Enzymol. 1992;213:403– 420.
3. Bhattacherjee P, Williams RN, Eakins KE. An evaluation of ocular
25. Di Mascio P, Kaiser S, Sies H. Lycopene as the most efficient
inflammation following the injection of bacterial endotoxin into
biological carotenoid singlet oxygen quencher. Arch Biochem
the rat foot pad. Invest Ophthalmol Vis Sci. 1983;24:196 –202.
4. Baatz H, Tonessen B, Prada J, Pleyer U. Thalidomide inhibits
26. Lim BP, Nagao A, Terao J, Tanaka K, Suzuki T, Takama K. Antioxidant
leukocyte-endothelium interaction in endotoxin-induced uveitis.
activity of xanthophylls on peroxyl radical-mediated phospholipid
Ophthalmic Res. 2001;33:256 –263.
peroxidation. Biochim Biophys Acta. 1992;1126:178 –184.
5. Ohta K, Nakayama K, Kurokawa T, Kikuchi T, Yoshimura N.
27. Shibata A, Kiba Y, Akati N, Fukuzawa K, Terada H. Molecular
Inhibitory effects of pyrrolidine dithiocarbamate on endotoxin-
characteristics of astaxanthin and beta-carotene in the phospho-
induced uveitis in Lewis rats. Invest Ophthalmol Vis Sci. 2002;43:
lipid monolayer and their distributions in the phospholipid bilayer. Chem Phys Lipids. 2001;113:11–22.
6. Rosenbaum JT, McDevitt HO, Guss RB, Egbert PR. Endotoxin-
28. Eigler A, Moeller J, Endres S. Exogenous and endogenous nitric oxide
induced uveitis in rats as a model for human disease. Nature.
attenuates tumor necrosis factor synthesis in the murine macrophage
cell line RAW 264.7. J Immunol. 1995;154:4048 – 4054. Anti-inflammatory Effect of Astaxanthin
29. Friden BE, Runesson E, Hahlin M, Brannstrom M. Evidence for
32. Eisenhut T, Sinha B, Grottrup-Wolfers E, Semmler J, Siess W,
nitric oxide acting as a luteolytic factor in the human corpus
Endres S. Prostacyclin analogs suppress the synthesis of
luteum. Mol Hum Reprod. 2000;6:397– 403.
tumor necrosis factor-alpha in LPS-stimulated human peripheral
30. Salvemini D, Misko TP, Masferrer JL, Seibert K, Currie MG, Needle-
blood mononuclear cells. Immunopharmacology. 1993;26:
man P. Nitric oxide activates cyclooxygenase enzymes. Proc NatlAcad Sci USA. 1993;90:7240 –7244.
33. Spengler RN, Spengler ML, Lincoln P, Remick DG, Strieter RM,
31. Endres S, Fulle HJ, Sinha B, et al. Cyclic nucleotides differentially
Kunkel SL. Dynamics of dibutyryl cyclic AMP- and prostaglandin
regulate the synthesis of tumour necrosis factor-alpha and interleu-
E2-mediated suppression of lipopolysaccharide-induced tumor ne-
kin-1 beta by human mononuclear cells. Immunology. 1991;72:56 –
crosis factor alpha gene expression. Infect Immun. 1989;57:2837–
SLEEP APNEA SYNDROMES The mysteries of sleep have intrigued man for centuries, it has only been within last three decades that we have begun to understand physiology and physiopatology of this state that occupies a considerable portion of our lives. It was developped a better understanding of specific sleep disorders and their realationship to common medical problems seen during wakefulnes
UPDATE: TOPROL-XL® (metoprolol succinate) now generally available after AstraZeneca delivers significant supply across the US in response to generic shortage Dear Doctor, We are pleased to provide you with follow-up information to our letter of last month regarding the availability of TOPROL-XL. As you know, after published reportsi,ii that two generic manufacturers had reca
 Anti-inflammatory Effect of Astaxanthin
Anti-inflammatory Effect of Astaxanthin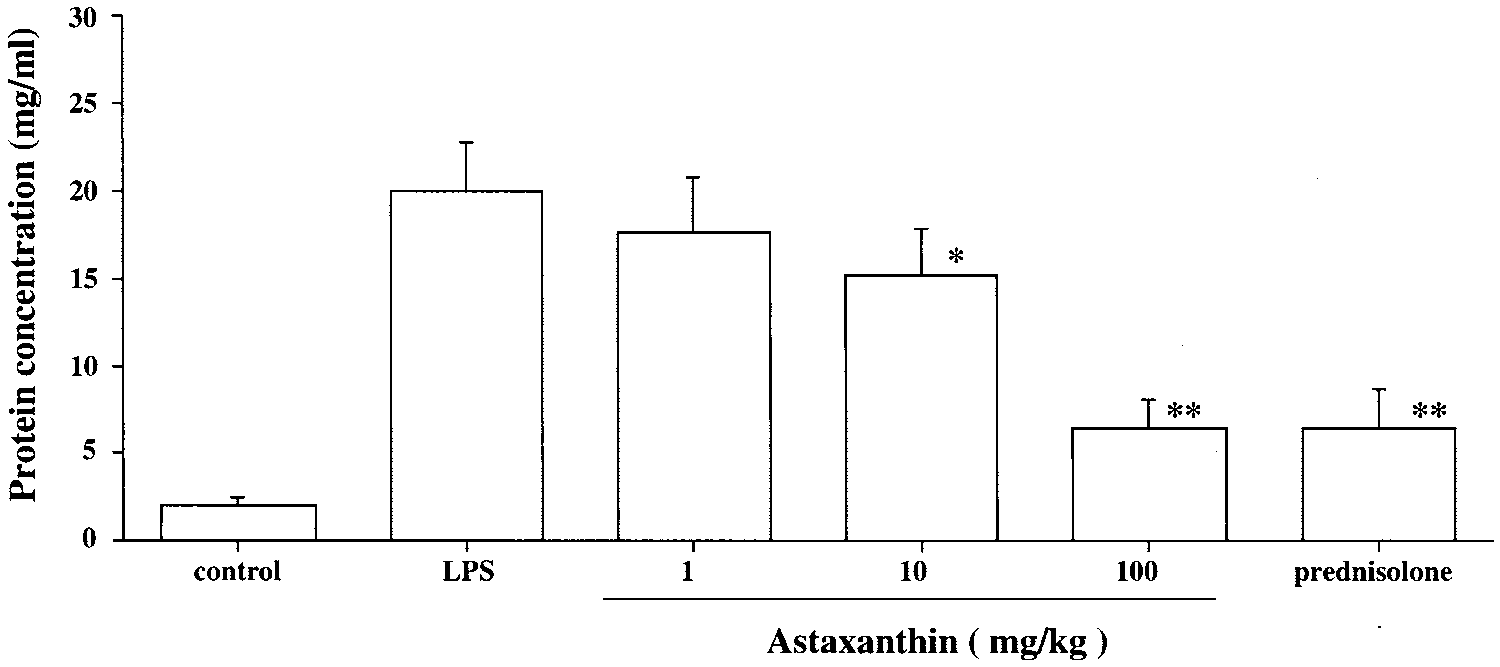
 Ohgami et al.
Ohgami et al.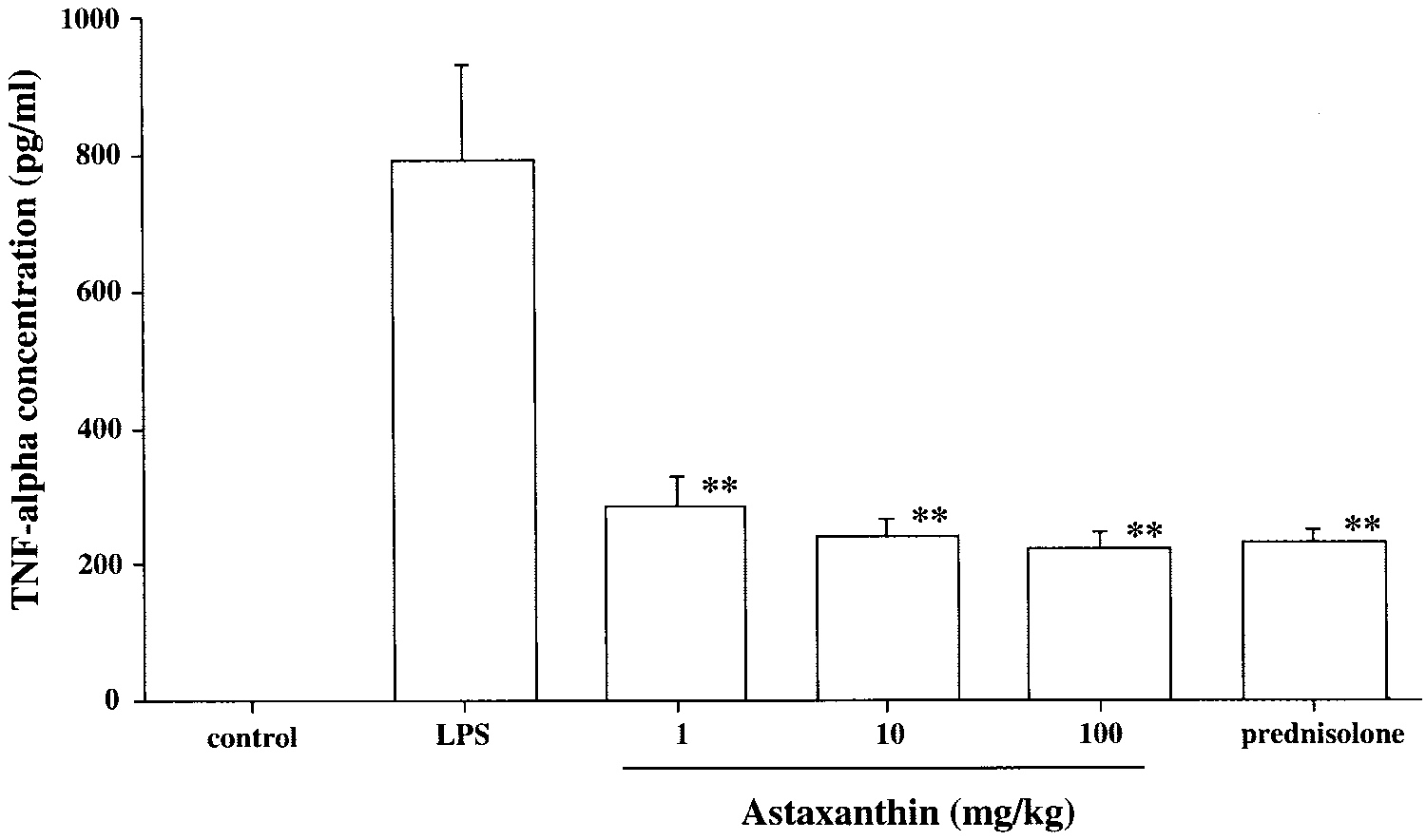
 Anti-inflammatory Effect of Astaxanthin
Anti-inflammatory Effect of Astaxanthin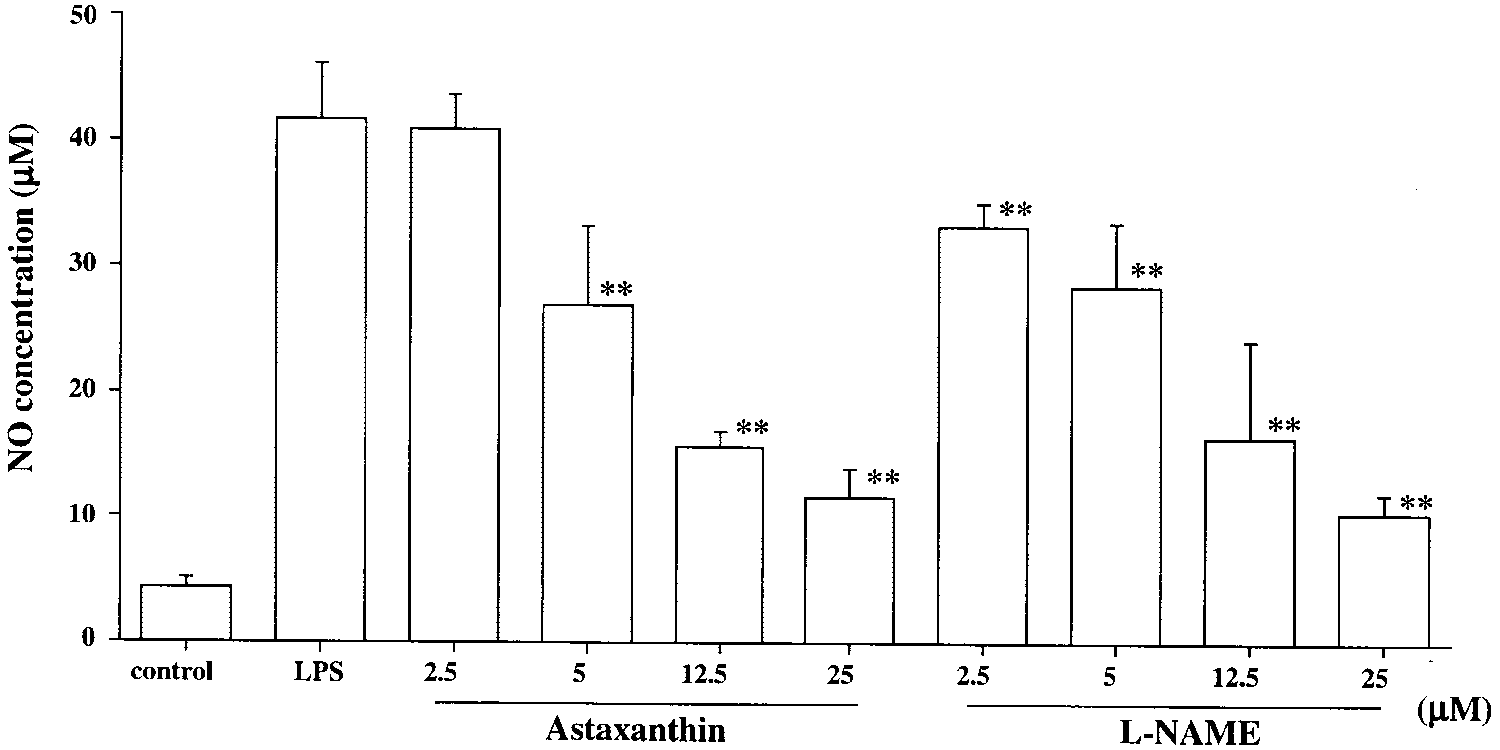
 Ohgami et al.
Ohgami et al.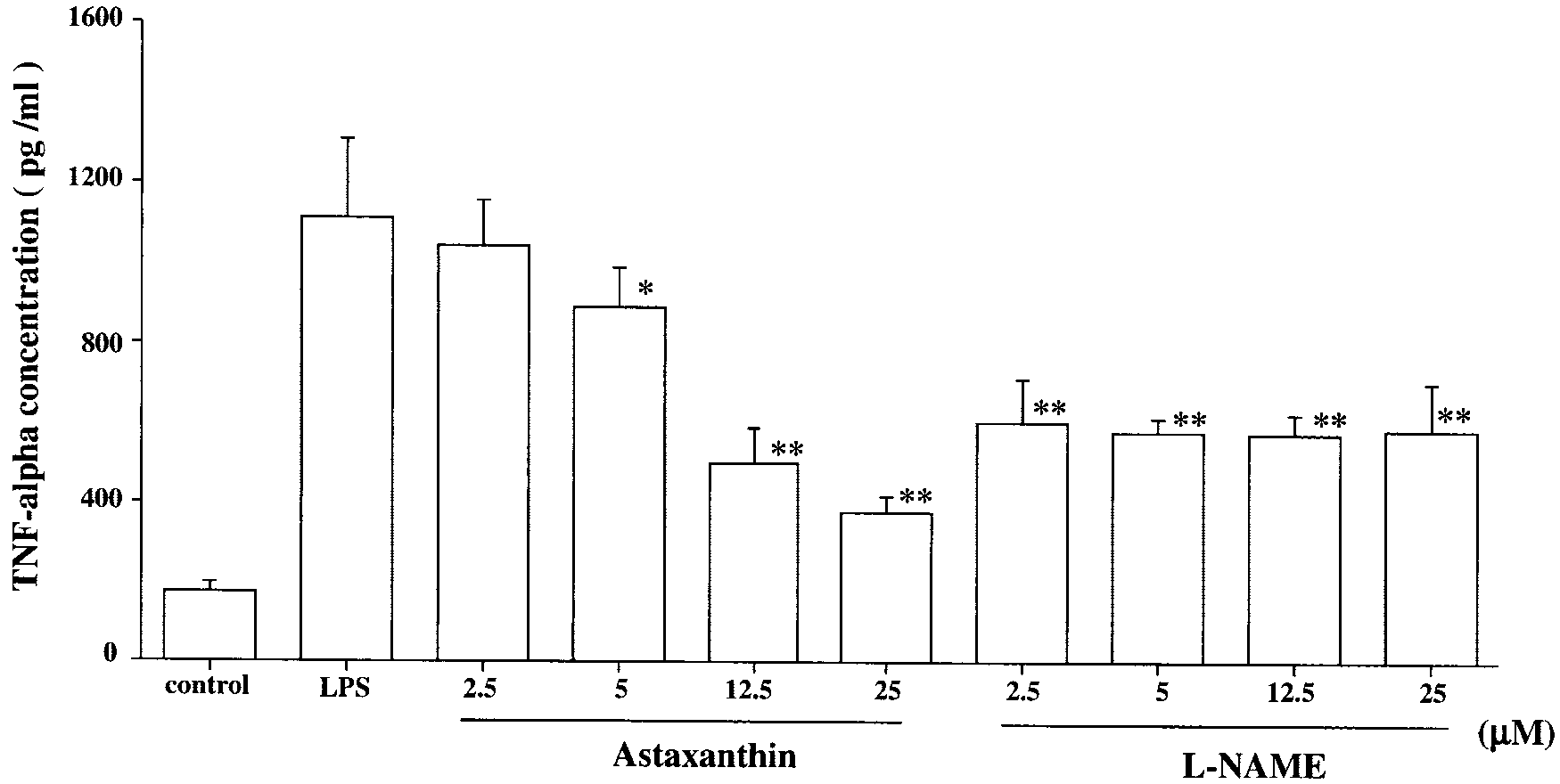
 Anti-inflammatory Effect of Astaxanthin
Anti-inflammatory Effect of Astaxanthin