Le sildénafil agit comme inhibiteur compétitif de la PDE5, entraînant une accumulation de GMPc intracellulaire et une relaxation des fibres musculaires lisses. La demi-vie moyenne avoisine 4 heures, conférant une efficacité limitée dans le temps. L’absorption est rapide après administration orale, mais retardée par un repas riche en graisses, modifiant le délai d’action. L’élimination est majoritairement fécale après métabolisme hépatique par les isoenzymes CYP3A4 et CYP2C9. Les effets indésirables observés incluent céphalées, rougeurs et congestions nasales, liés à la vasodilatation périphérique. Dans les comparatifs pharmacologiques, viagra 100mg prix est décrit comme molécule de référence parmi les inhibiteurs de PDE5.
Rogerslab.org.uk
Neuropsychopharmacology (2006) 31, 1075–1084& 2006 Nature Publishing Group
Effects of Tryptophan Depletion on the Performance of anIterated Prisoner’s Dilemma Game in Healthy Adults
Richard M Wood1, James K Rilling2, Alan G Sanfey2, Zubin Bhagwagar3 and Robert D Rogers*,1
University Department of Psychiatry, University of Oxford, Oxford, UK; 2Departments of Anthropology, Psychiatry and Behavioral Sciences,
Emory University, Atlanta, GA, USA; 3Department of Psychiatry, Yale University, New Haven, CT, USA
Adaptive social behavior often necessitates choosing to cooperate with others for long-term gains at the expense of noncooperative
behaviors giving larger immediate gains. Although little is know about the neural substrates that support cooperative over
noncooperative behaviors, recent research has shown that mutually cooperative behavior in the context of a mixed-motive game, the
Prisoner’s Dilemma (PD), is associated with increased neural activity within reinforcement circuitry. Other research attests to a role for
serotonin in the modulation of social behavior and in reward processing. In this study, we used a within-subject, crossover, double-blind
design to investigate performance of an iterated, sequential PD game for monetary reward by healthy human adult participants following
ingestion of an amino-acid drink that either did (T + ) or did not (TÀ) contain l-tryptophan. Tryptophan depletion produced significant
reductions in the level of cooperation shown by participants when playing the game on the first, but not the second, study days. This
effect was accompanied by a significantly diminished probability of cooperative responding given previous mutually cooperative behavior.
These data suggest that serotonin plays a significant role in the acquisition of socially cooperative behavior in human adult participants,
and suggest novel hypotheses concerning the serotonergic modulation of reward information in socially cooperative behavior in both
Neuropsychopharmacology (2006) 31, 1075–1084. doi:10.1038/sj.npp.1300932; published online 11 January 2006
Keywords: Prisoner’s Dilemma; serotonin; reciprocal cooperation; social function; aggression; reward
evolutionary contexts (Axelrod, 1984; Axelrod and Hamil-ton, 1981), individual differences in competitive and
Mutually cooperative behavior among individuals is an
exploitative strategies during reciprocal interactions (Pruitt
integral part of greater-ape society, yet there has been
and Kimmel, 1977), and to identify factors that promote or
relatively limited research into the nature of the neural
inhibit cooperation in diverse experimental and naturalistic
substrates supporting social cooperation in human and
settings (Axelrod, 1984; Boone et al, 2002; Sheldon, 1999).
non-human species. The Prisoner’s Dilemma (PD) offers a
Rilling et al (2002) used functional magnetic resonance
tractable laboratory measure of social cooperation based
imaging (fMRI) to investigate regional changes in blood-
upon reciprocal altruism which can be used to advance this
oxygenation-level-dependent response (BOLD) associated
area experimentally (Trivers, 1971, 1985): two players make
with socially cooperative behavior in an iterated PD game
choices associated with certain pay-offs (represented by a
(viz., a game which is played several consecutive times or
‘pay-off matrix’; see below). The pattern of choices made
‘rounds’ with the same partner). Mutually cooperative
can lead to an equal sharing or an unequal sharing of the
outcomes (in which both players chose to cooperate: CC)
pay-offs (favoring one player or the other), thereby allowing
were associated with increased BOLD within the orbito-
players to display cooperation or selfishness. This model
frontal cortex compared to mixed outcomes (in which one
has been used extensively by researchers from many
of the two players chose to cooperate, while the other chose
disciplines to explore the emergence of cooperation in
to defect: CD or DC) and mutual defection outcomes (inwhich both players chose to defect: DD). Although this was
*Correspondence: Dr RD Rogers, Department of Psychiatry, Uni-
the case when players believed they were playing with a
versity of Oxford, Warneford Hospital, Oxford, Oxfordshire OX3 7JX,
human partner or a computer, mutual cooperation was
UK, Tel: + 44 1865 226 399, Fax: + 44 1865 793 101,
associated with greater activation in the rostral anterior
cingulate cortex and anteroventral striatum specifically
Received 23 December 2004; revised 21 July 2005; accepted 22August 2005
when participants believed they were playing with a human
Online publication: 9 September 2005 at http://www.acnp.org/citations/
partner. These neural systems receive dopamine projec-
tions from the midbrain and are known to be involved
Effects of tryptophan depletion on PD game performance
in reward and the processing of motivationally significant
Chronic citalopram was associated with increased choices
information (Robbins and Everitt, 1996), suggesting that
of the more generous pay-offs and increased affiliative
socially cooperative behavior is supported by activity within
messages sent to the playing partner (Tse and Bond, 2002a).
reinforcement pathways (Moskowitz and Cote, 1995). A
In a similar study, reboxetine increased cooperation in the
follow-up study using a one-shot PD paradigm in which
PD game, while citalopram sustained emotional expression
participants played a new partner on each round con-
of speech when speaking to a socially withdrawn partner
firmed this hypothesis by demonstrating increased BOLD
(Tse and Bond, 2002b). Summarizing the above results,
within subcallosal anterior cingulate cortex, anteroventral
increasing serotonin activity appears to change indirect
striatum, and medial orbitofrontal cortex with mutually
measures of social function in healthy human adults, and
cooperative outcomes (Rilling et al, 2004).
may increase affiliative gestures in dyadic interactions.
Observations in several species indicate a role for
However, there is little information about the effects of
serotonin in the modulation of prosocial behavior. Inves-
reducing serotonin on cooperative behavior in mixed-
tigation of both peripheral and central indices of serotonin
motive games, such as a true iterated PD, where the gains
function have shown negative associations with violent and
and losses for one player depend upon effective reciprocal
aggressive behavior in rhesus monkeys (Higley et al, 1992,
exchanges with a social partner. We sought to investigate
1996) and human clinical samples (Brown et al, 1979;
Virkkunen et al, 1994), as well as positive associations with
We used rapid tryptophan depletion to restrict the
socially affiliative behaviors (such as grooming and
availability of l-tryptophan for serotonin synthesis in
approach) (Raleigh et al, 1981). Pharmacological interven-
healthy human participants in order to explore subsequent
tions that increase serotonergic activity also attest to a
changes on performance of an iterated PD game for
prominent role in important aspects of social behavior and
monetary reward. This methodology has shown to be an
aggression (for a discussion, see Fuller, 1996). The selective
effective way of temporarily reducing serotonin activity in
serotonin reuptake inhibitor (SSRI), fluoxetine, increases
both animal and humans (Biggio et al, 1974; Carpenter et al,
affiliative behaviors in rodents (Knutson and Panksepp,
1998), with evidence of cognitive effects in healthy
1996), while each of fluoxetine, quipazine (a 5-HT1 and
participants without a history of mood disorder (Park
5-HT2 receptor agonist), and the amino-acid serotonin
et al, 1994; Rogers et al, 1999; Moore et al, 2000). The
precursor, l-tryptophan, have been found to increase
starting point for our study was the accumulating evidence
affiliative behaviors and decrease nonsocial behaviors (such
that serotonin plays a role in modulating the various aspects
as vigilance and avoidance) in primates (Raleigh et al,
of reward processing (Aronson et al, 1995; Redgrave and
1985). Additional information about these effects has been
Horrell, 1976; Sasaki-Adams and Kelley, 2001) mediated by
provided by observations that fluoxetine and l-tryptophan
circuitry encompassing orbitofrontal and rostral cingulate
increase the social dominance of subdominant vervet males
cortices, and ventral striatum (Robbins et al, 1989). We
in the absence of the alpha male, while interventions that
wished to test the hypothesis that reducing serotonergic
antagonise serotonin (eg the 5-HT2C receptor antagonist,
modulation in healthy participants would reduce the
cyproheptadine, and chronic treatment with fenfluramine)
number of cooperations and increase the number of
prevent the emergence of dominance under these condi-
defections in an iterated PD game, perhaps by interfering
with activity within reinforcement-related circuitry asso-
In humans, major depressive disorder is widely accepted
ciated with mutually cooperative behavior.
as involving altered serotonin function and is characterizedby relatively poor social functioning (Fossi et al, 1984;Weissman, 2000). Consistent with this finding, antidepres-
sant medications that enhance serotonergic function havebeen shown to increase affiliative behaviors, including
The study was approved by the Oxfordshire Psychiatry
verbal and nonverbal positive communication in depressed
Research Ethics Committee, UK. All participants gave
patients, sometimes independently of effects on depressed
mood (Dubini and Polin, 1997), and can reduce hostilesentiment and violent outbursts in impulsive clinical
samples (Coccaro and Kavoussi, 1997).
Despite this, experimental investigation of the role of
A total of 24 healthy adults (12 males; 12 females)
serotonin in the social behavior of human participants has
participated. Previous research has shown that tryptophan
been limited. Chronic l-tryptophan intake has been found to
depletion in participants with a history of mood disorder
both increase dominant behaviors, and decrease quarrel-
can reinstate dysphoria (Moore et al, 2000). Therefore,
someness, in everyday interactions (Moskowitz et al, 2001),
participants were carefully examined by an experienced
while the SSRI, paroxetine, has been found to increase
psychiatrist (ZB) to ensure that none of the following
affiliative gestures towards a partner during completion of a
exclusion criteria were met: (i) major physical illness, (ii)
puzzle task after 1 but not 4 weeks of treatment (Knutson
current or previous DSM-IV major mood disorder as
et al, 1998). More formally, Tse and Bond (2002a, b) have
assessed by a SCID-I interview (First et al, 2002), and (iii)
examined the effects of chronic treatment with the SSRI,
current or previous substance abuse. Participants with a
citalopram, and the selective noradrenergic reuptake
family history of mood disorder were not excluded. There
inhibitor, reboxetine, on the performance of an amended
was no restriction on the phase of menstrual cycle in female
PD game involving the division of pay-offs between the two
participants. Verbal IQ was estimated with the National
players and communication with the playing partner.
Effects of tryptophan depletion on PD game performanceRM Wood et al
game, each participant was introduced to a gender-matched‘partner’Fin reality, an experimenter-confederate. It was
The study consisted of a within-subject, crossover, double-
explained to the participant that his/her computer was
blind design. Each participant ingested an amino-acid drink
networked to a computer in another room in which their
on 2 study days, separated by not less than 1 week. One
partner would play. The experimenter left to take the
drink contained l-tryptophan (T + ), the other drink did not
confederate to their room, returning a few minutes later to
(TÀ). The order of these treatments was counterbalanced
talk the participant through the PD game instructions.
across the participant group. A total of 12 participants under-
In fact, the confederate’s choices were made by the
went the T + treatment first and the TÀ treatment second
computer, running a predetermined strategy of tit-for-tat
(six male and six female) (T + /TÀ). A total of 12 participants
underwent these treatments in the reverse order (six male
On each round of the PD game, the participant viewed a
2 Â 2 matrix that displayed the pay-off matrix for that round(see Figure 1). The four squares of this pay-off matrix
defined four possible outcomes: both players cooperate(CC); the participant cooperates but the partner defects
Amino acids were supplied by SHS International Ltd.
(CD), the participant defects but the partner cooperates
The amounts of each for male and female participants,
(DC), and both players defect (DD). The payoffs for the
respectively, were l-alanine (5.5 g; 4.58 g), l-arganine (4.9 g;
outcomes were constructed such that DC4CC4DD4CD,
4.08 g), l-cystine (2.7 g; 2.25 g), glycine (3.2 g; 2.67 g),
and CC4(CD + DC)/2 (2). Specifically, if the participant
l-isoleucine (8.0 g; 6.67 g), l-leucine (13.5 g; 11.25 g), l-lysine
and partner cooperated (CC), both would receive 20p. If
monohydrochloride (11.0 g; 9.17 g), l-methionine (3.0 g;
the participant defected while the partner cooperated (DC),
2.5 g), histidine (3.2 g; 2.67 g); l-phenylalinine (5.7 g; 4.75 g),
the participant would earn 30p, and the partner nothing;
l-proline (12.2 g; 10.17 g), l-serine (6.9 g; 5.75 g), l-threonine
and vice versa (CD). If both players defected (DD), both
(6.5 g; 5.42 g), l-tyrosine (6.9 g; 5.75 g), l-valine (8.9 g; 7.42 g).
The T + drink contained l-tryptophan (2.3 g; 1.92 g). The
The participant’s partner always chose first (ie the game
unpleasant taste of the drinks was masked using 5 g flavor
was sequential). If the partner cooperated, the upper row of
sachets. Each sachet consisted of citric acid (or malic acid),
the matrix was highlighted in yellow. If the partner defected,
flavorings (lemon-and-lime, cherry-and-vanilla, or grape
the lower row was highlighted in yellow. After this, the
fruit), and artificial sweetener. Each sachet contained
participant responded with his own choice. If he/she
approximately 15 calories, and 1.3 g of carbohydrate.
cooperated, the left column was highlighted; if he/shedefected, the right column was highlighted. Consequently,
the combination of the partner’s and the participant’schoices specified the outcome of the round. The participant
Participants followed a low-protein diet (less than 20 g) the
pressed the ‘C’ key on the computer keyboard to indicate a
day before each study day. Participants attended the
cooperative response, and the ‘D’ to indicate a defection
laboratory at 0830 hours. Blood samples were taken to
response. Before starting, four training trials demonstrated
obtain baseline levels of plasma total l-tryptophan. Partici-
the four possible choice combinations; the participant was
pants then drank the amino-acid drink over a 30-min
also shown a table showing the total earnings arising
period. Some participants reported transitory nausea
through the four possible choice outcomes over the course
and tiredness. Participants were given a low-protein lunch
(less than 2 g). At 5 h after consuming the amino-acid drink( + 5 h), a second blood sample was taken. Participantsthen completed the PD game.
Participants completed self-report measures of state
positive and negative affect (PANAS; Watson et al, 1988)and validated visual analog scales of aggression (VAS; Bondand Lader, 1974) at baseline and + 5 h. Participants alsocompleted psychometric assessments of trait aggressionusing the Buss–Perry Aggression Questionnaire (AQ; Bussand Perry, 1992). The AQ provides subscales for physicaland verbal aggression, hostility, and anger. The VAS wasused to assess effects of tryptophan depletion on partici-pants’ mood, while the AQ was used to assess whethereffects on social cooperation involved differences in traitaggression.
The iterated, sequential PD game. The iterated, sequentialPD used in the study was adapted from that reported by
Payoff matrix for the four outcomes in the iterated, sequential
Prisoner’s Dilemma (PD) game. The participant’s choices (‘You’) are listed
Rilling et al (2002). The game was presented on a computer
atop columns and the partner’s choices (‘partner’; in fact, a computer
that had connecting cables going into the laboratory wall,
program playing tit-for-tat) are listed aside the rows. The payoffs for each
facilitating the deception that the computer was networked
player, depending upon both players’ choices, are shown within each
to a different computer in another room. Before playing the
square; green ¼ participant’s and pink ¼ partner’s).
Effects of tryptophan depletion on PD game performance
Table 1 Monetary Payoffs for Four Different Strategies Over the
tion of the data from the PD games.) Where the effects of
Course of the Iterated, Sequential Prisoner’s Dilemma (PD) Game
the TÀ and T + treatments were different on the first and
second study days, as indicated by significant two-wayinteractions between treatment and treatment order,
dependent measures were subject to supplementary ANO-VAs for each study day separately (to investigate simple
Both you and your partner cooperate each round
interaction effects), with gender and treatment (T + vs TÀ)
Your partner cooperates and you defect each round
as two between-subject factors and game as a single within-
Your partner defects and you cooperate each round
Both you and your partner defect each round
‘You’ ¼ participant; ‘Partner’ ¼ computer program playing tit-for-tat.
Participants’ mean age and verbal IQ were 27.0471.49 (SE)
Participants played two separate PD games, each consist-
and 113.8771.43. There were no significant differences
ing of 20 rounds. The procedure contained two manipula-
between the age or estimated IQs of those participants who
tions. First, the computer started the first game by choosing
completed the T + /TÀ treatment order (27.8972.26 years
to cooperate but started the second game by choosing to
and 11271.98) and those who completed the TÀ/T + order
defect. Second, after its first choice in both games, the
(26.2072.00 and 11671.98) (Fs(1, 20)o2.35). Similarly,
computer followed a strict tit-for-tat strategy, always
the two groups were matched for trait positive affect
mimicking the responses of the participants on the previous
(36.8371.52 vs 33.9271.26; F(1, 20) ¼ 2.45), trait negative
round. Participants were thoroughly debriefed about the
affect (15.3371.80 vs 13.2570.70; F(1, 20) ¼ 1.14), total self-
deception at the end of the study; all indicated that they had
report aggression (6272.45 vs 66.1874.14; Fo1.00),
believed that they had been playing the PD game with a real
physical aggression (14.2570.90 vs 17.0971.88; F(1, 15) ¼
human partner (ie the confederate) rather than a computer
1.31), verbal aggression (15.0070.46 vs 16.0970.63;
F(1, 15) ¼ 1.61), anger (14.3870.98 vs 16.3671.27; F(1, 15) ¼1.15), and hostility (18.3871.21 vs 16.6471.30; F(1, 15) ¼1.04). (For technical reasons, scores from the AQ (Buss and
Perry, 1992) were unavailable for four participants who
The dependent measures for the PD game were as follows:
completed the T + /TÀ treatment order and for one
(i) the proportion of rounds on which participants chose
participant who completed the TÀ/T + treatment order.)
to cooperate (‘proportionate choice’); (ii) the deliberationtime (in milliseconds) taken to make a cooperative or
a defection choice; (iii) the ‘conditional probability’ ofcooperating specified as the proportion of rounds on which
As expected (Moore et al, 2000), total plasma tryptophan
participants cooperated following each of the four possible
was reduced between baseline and + 5 h after the TÀ
outcomes (CC, CD, DC, DD) on the immediately previous
treatment (12.4470.68 vs 1.9270.27 mg/ml) but increased
after the T + treatment (13.0170.51 vs 30.2772.31 mg/ml)
Differences in age, estimated verbal IQ, and psychometric
(F(1, 12) ¼ 123.43, po0.0001). Analysis of simple effects
measures of self-report trait affect and aggression were
confirmed that the difference in plasma tryptophan follow-
analyzed by analysis of variance (ANOVA) with treatment
ing the TÀ and T + treatments was significant at + 5 h
order across the two study days (T + /TÀ and TÀ/T + ) and
Proportionate choice of cooperative responses were
analyzed with repeated measures ANOVAs with gender,treatment order as between-subject factors, and treatment
Treatment with the TÀ drink produced different kinds of
(T + vs TÀ) and game (1 vs 2) as within-subject factors. The
effects on cooperative behavior on the 2 study days as
reaction times (ms) for cooperative and defection responses
evidenced by a significant two-way interaction between
were averaged together to give mean deliberation times and
treatment and treatment order (F(1, 20) ¼ 4.68, po0.05).
submitted to the same analysis as the proportionate choice
Statistical analysis of each study day separately demon-
data. Plasma tryptophan, state positive affect, state negative
strated that the TÀ treatment was associated with
affect (PANAS), and state aggression (VAS) were analyzed
significantly fewer cooperative responses than the T +
with repeated measures ANOVAs with gender and treat-
treatment on day 1 (F(1, 20) ¼ 6.67, po0.05) (see Figure 2),
ment order as between subject-factors and treatment (T +
but a slightly increased number of such responses on day 2
vs TÀ) and time (baseline vs + 5 h) as within-subject
(F(1, 20) ¼ 2.63) (Figure 2). Pair-wise tests showed that
factors. (The 13 VAS subscales were included in order to
participants who underwent the TÀ treatment on day 1
assess whether the performance of the PD games following
showed a significant increase in cooperative responses after
tryptophan depletion might be attributable to short-term
the T + treatment on day 2 (0.5070.07 to 0.6470.08;
changes in aggression. We did not correct for multiple
t (two-tailed) ¼ 2.26, df ¼ 11, po0.05), while participants
comparisons when analyzing this data in order to highlight
who underwent the T + treatment on day 1 showed
any such changes that might be relevant to the interpreta-
no significant change in cooperative responses after the
Effects of tryptophan depletion on PD game performanceRM Wood et al
TÀ treatment on day 2 (0.7370.05 to 0.8070.04; t (two-
However, analysis of each study day separately showed that
tailed) ¼ À0.98, df ¼ 11, po0.35).
the deliberation times following the TÀ and T + treatments
Overall, there was a trend for all participants to make
did not differ significantly on day 1 (32497421 ms vs
more cooperative responses on the first game of 20 rounds
29957387 ms, respectively) (Fo1.00) or day 2 (23377
than the second game of 20 rounds (0.7470.05 vs
279 ms vs 20427208 ms; Fo1.00). Mean deliberation times
0.5970.06), (F(1, 20) ¼ 3.49, p ¼ 0.08). There were no
in the first game of 20 rounds were not significantly
significant differences between male and female participants
different from those in the second game (28607268 vs
in the proportion of cooperative responses (0.6870.06 vs
24527190 ms) (F(1, 20) ¼ 2.94). Male participants did not
0.6570.06, respectively; Fo1.0). There were no significant
significantly differ from female participants in their
interactions involving treatment, gender, and game.
deliberation times (27567359 vs 25577197 ms) (Fo1.00).
Treatment with the TÀ drink affected participants’
There were no other significant interactions involving
deliberation times differently on the first and second study
days, as evidenced by a two-way interaction between
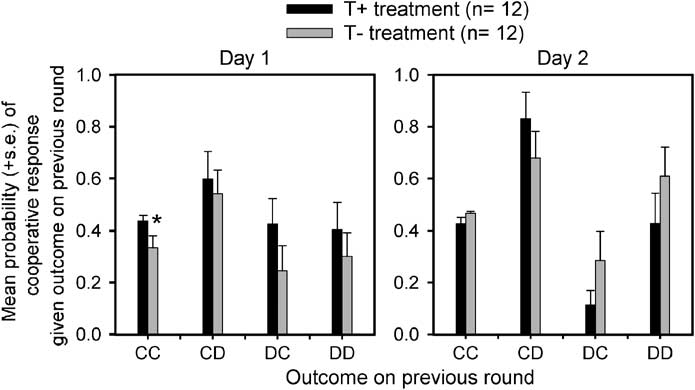
Conditional probabilities of cooperative responses were
treatment and treatment order (F(1, 20) ¼ 15.42, po0.005).
defined as the proportion of trials on which participantsmade a cooperative choice given one of the four outcomes(CC, CD, DC, and DD) on the previous round of the game. On the first study day, the TÀ participants exhibited asignificantly reduced conditional probability of a coopera-tive response given a mutual cooperative outcome on theprevious round compared to the T + treatment (seeFigure 3; F(1, 22) ¼ 4.35, po0.05); no other differencesin the conditional probability of cooperative responseswere significant (all Fsp1.07). On the second day, theTÀ treatment produced a slightly increased conditionalprobability of a cooperative response given a mutual co-operative outcome on the previous round compared to theT + treatment; this effect was not significant (Figure 3;F(1, 22) ¼ 2.42).
Overall, state positive affect was significantly reducedbetween baseline and + 5 h (F(1, 20) ¼ 15.88, po0.005),but no more so after the TÀ treatment (31.2971.42 vs
Mean proportion of cooperative choices of healthy participants
28.9671.47) compared to the T + treatment (30.4271.61 vs
who drank an amino-acid drink containing l-tryptophan (T + ) treatment
and participants who drank an amino-acid drink not containing l-tryptophan
71.54) (Fo1.00). State negative affect did not change
(TÀ treatment) on the first and second study days. * ¼ F(1, 20) ¼ 6.67,
markedly between baseline and + 5 h (F(1, 20) ¼ 2.29) after
either the TÀ treatment (12.7170.94 vs 12.4671.01) or T +
The conditional probability of making a cooperative choice given the four possible outcomes of the previous round in participants who drank an
amino-acid drink containing l-tryptophan (T+) and in participants who drank an amino-acid drink not containing l-tryptophan (TÀ) on the first and secondstudy days (CC ¼ participant cooperates–partner cooperates, CD ¼ participant cooperates–partner defects, DC ¼ participant defects–partner cooperates,DD ¼ participant defects–partner defects). * ¼ F(1, 21) ¼ 4.352, po0.05.
Effects of tryptophan depletion on PD game performance
treatment (13.5471.02 vs 12.6370.69) (Fso1.00). There
TÀ compared to the T + treatment on either study day (see
were no significant differences in the state positive or
Table 2). Moreover, the reduced cooperation of those
negative affect of the T + and TÀ participants at baseline
participants tested after the TÀ treatment on study day 1
or + 5 h on either study day 1 or day 2 (all Fso1.00;
cannot be attributed to an increase in state aggression in
see Table 2). Entering state positive and negative affect
comparison with those tested after the T + treatment. These
at + 5 h on day 1 as covariates did not abolish the
two groups of participants were matched on the multiple
significant reduction in cooperative responses associated
facets of aggression measured by the VAS (Bond and Lader,
with the TÀ compared to T + treatments (F(1, 18) ¼ 6.70,
1974) at baseline, and showed broadly similar changes
during the interval between baseline and playing the PDgame + 5 h later. At that time, 11 of the 13 VAS showedlower scores associated with the TÀ treatment compared to
the T + treatment and only two showed higher scores (see
Overall, self-reported annoyance was significantly decreased
Table 2). Indeed, the single significant treatment difference
after the TÀ compared to the T + treatment (21.7772.62 vs
(uncorrected for multiple comparisons) at the time of
26.0473.49) (F(1, 20) ¼ 4.88, po0.05); self-reported disgust
playing the PD game on day 1 indicated that participants
was also reduced after the TÀ treatment compared to the
tested after the TÀ treatment rated themselves as signifi-
T + treatment (21.8172.74 vs 29.1073.64) (F(1, 20) ¼ 5.48,
cantly less rebellious than those tested after the T +
po0.05). Comparisons of ratings at + 5 h on study day 1,
treatment. In summary, there is no evidence to suggest
taken just before playing the PD game, revealed that
that the reduced cooperation associated with the TÀ treat-
rebelliousness was significantly lower following the TÀ
ment on the first study day is explicable by differences in
state affect, or increased state aggressivity, consequent
state furiousness showed a similar trend (F(1, 20) ¼ 3.48,
to tryptophan depletion. Rather, the data indicate that
p ¼ 0.08). There were no other significant treatment-related
serotonin can affect social cooperative behavior indepen-
differences in state aggression ratings at baseline or + 5 h
dently of the gross changes in emotional state that might be
detectable by self-report measures of positive or negativeaffect including state aggression.
Similarly, those participants tested after the TÀ treatment
and those tested after the T + treatment on the first studyday also scored equivalently on trait positive and negative
The iterated PD has been used extensively by researchers in
affect, as well as on self-report trait physical aggression,
social psychology, economics, and game theory to model
trait verbal aggression, trait anger, and trait hostility.
social relationships involving reciprocal altruism (Axelrod,
Consequently, the reduced cooperation in the PD game
1984; Axelrod and Hamilton, 1981; Trivers, 1971, 1985).
associated with the TÀ treatment cannot be the result of
These results demonstrate that healthy adult participants
between-subject variation in personality traits reflecting
who ingested an amino-acid drink lacking l-tryptophan
stable aspects of emotional function and aggression. (The
(TÀ treatment) showed a pattern of reduced cooperative
two participant groups were also closely matched for age
responses on an iterated, sequential PD game compared
and estimated verbal IQ.) Accumulating data suggest that
to participants who drank a tryptophan-balanced drink
the effects of tryptophan depletion sometimes depend upon
(T + treatment) on the first, but not second, day of the
variation in associated traits linked to serotonergic func-
study (Figure 2). Additionally, the conditional probability
tion. Specifically, manipulation of l-tryptophan has been
of a cooperative response given a mutually cooperative
found to alter reactive aggression in participants with high,
outcome on the previous round of the game was similarly
but not low, trait aggression (Cleare and Bond, 1995; Bjork
and significantly diminished (Figure 3). Consequently,
et al, 2000) and in participants vulnerable to alcoholism
these findings suggest that temporary reduction of
(LeMarquand et al, 1999; Marsh et al, 2002). Such findings
central serotonin actitivity, achieved by manipulation
are consistent with evidence, cited above, that aggressive
of l-tryptophan, impairs the ability of healthy adults to
behavior depends upon serotonergic mechanisms in experi-
learn (or acquire) a pattern of cooperative behavior in the
mental, clinical, and naturalistic settings (Raleigh et al,
context of a validated laboratory measure of reciprocal
1991; Virkkunen et al, 1994; Giovanni et al, 2001 for
a review). However, the present data also demonstrate
We acknowledge that the above pattern of data may
an effect of tryptophan depletion on the acquisition of
reflect the influence of uncontrolled factors such as family
cooperative behavior in a laboratory measure of social
history of mood disorder and, in the female participants,
cooperation that appears largely independent of variation
phase of menstrual cycle that might have interacted with
in physical or verbal aspects of trait aggression, its
tryptophan depletion to compromise cooperative behavior
(common) emotional correlate (anger), or the possession
while playing the PD game. However, importantly, the
of critical social attitudes (as indexed by trait hostility).
reduced cooperative responses associated with the TÀ treat-
One notable feature of our results is that there was a
ment on the first study day are not attributable to, or
significant interaction between treatment and treatment
secondary to, temporary changes in mood. Consistent with
order such that tryptophan depletion significantly affected
previous studies (Moore et al, 2000), our participants, who
performance of the PD game on the first but not the second
did not include any with a history of mood disorder, did
study day. Similar interactions have been reported fre-
not report significantly greater reductions in state posi-
quently in pharmacological studies of cognitive function
tive affect, or increases in state negative affect, after the
that have used within-subject, crossover designs, including
Effects of tryptophan depletion on PD game performanceRM Wood et al
Table 2 Subjective Effects for Participants Who Drank an Amino-Acid Drink Containing l-Tryptophan (T+) and Participants Who Drankan Amino-Acid Drink not Containing l-Tryptophan (TÀ) on Study Day 1 and Day 2
State positive and negative affect (PANAS). Visual analog scales for state aggression (VAS). * ¼ F(1,20) ¼ 5.83, po0.05.
Effects of tryptophan depletion on PD game performance
some involving tryptophan depletion (Park et al, 1994).
cingulate cortex and anteroventral striatum compared to all
These effects reflect the fact that drug treatments can have
other outcomes of the game (ie mixed cooperation/
different effects depending upon whether tasks are com-
defection and mutual defection). That data suggest that
pleted for the first or second time and are particularly
mutual cooperation activates neural systems of the fore-
marked where the tasks involve significant learning (Coull
brain, perhaps supporting the development of social
et al, 1995; Elliott et al, 1997). In the present study, analysis
reciprocity over repeated encounters (Rilling et al, 2002,
of the simple effects indicated that tryptophan depletion
2004). Evidence has continued to accumulate suggesting
reduced participants’ cooperative behavior on the first
that serotonin plays a significant role in modulating the
study day by approximately 31%. This impression was
reinforcement processes mediated by this circuitry and its
reinforced by pair-wise comparisons indicating that parti-
mid-brain dopaminergic innervation. This evidence in-
cipants who underwent the TÀ treatment on study day 1,
cludes demonstrations that serotonergic activity enhances
and who showed relatively reduced number of cooperative
intracranial self-stimulation (Redgrave and Horrell, 1976)
responses, significantly increased their cooperative re-
and that SSRI treatment decreases self-stimulation thresh-
sponses after the T + treatment on study day 2, while
olds (Harrison and Markou, 2001). Additionally, potentiat-
participants who underwent the T + treatment on day 1
ing and diminishing serotonergic activity has been found to
showed similarly high levels of cooperation on day 2.
increase and decrease the reinforcing effects of cocaine,
Evidence indicates that the tit-for-tat strategy followed by
respectively (Aronson et al, 1995; Sasaki-Adams and Kelley,
the fictional partner of the PD game in the present study
2001). Finally, Rogers et al (2003) recently demonstrated
(actually a prepared computer program) is highly effective
that tryptophan depletion altered healthy participants’
in eliciting cooperation from other playing partners
processing of prospective gains, but not prospective
(Sheldon, 1999). Consequently, these data suggest that
losses or probability cues, when choosing between binary-
temporary disruption of serotonin impairs the acquisition
outcome gambles, suggesting that serotonin can, under
of cooperative behaviors even when the strategies of social
appropriate conditions, modulate risky choice through the
partners are geared to elicit reciprocal cooperation.
control of attention towards appetitive signals. Therefore,
Serotonin has also been shown to influence learning
our finding that tryptophan depletion reduces cooperative
mechanisms that may mediate acquisition of socially
choices on an iterated PD game of the kind employed in the
cooperative behavior (Fletcher et al, 1999; Sasaki-Adams
present study suggests that serotonin modulates activity
and Kelley, 2001) and which depend upon much the same
within circuitry encompassing the rostro-cingulate cortex
neural circuitry as performance of an iterated PD game
and the striatum that mediates the reinforcing effects of
(Rilling et al, 2002). Serotonergic depletions within the
cooperative behavior (Rilling et al, 2002). This hypothesis
forebrain of monkeys (Clarke et al, 2004), and tryptophan
may provide a basis for understanding why major
depletion in humans (Park et al, 1994; Rogers et al, 1999),
depressive disorder is associated with social withdrawal
impair the ability to learn and relearn changing stimulus-
(see Fossi et al, 1984; Weissman, 2000) by positing a failure
reinforcement associations, probably reflecting altered
of serotonergic innervation to modulate appropriately the
neuromodulation of circuitry encompassing the orbito-
reinforcement value of social interactions (Libet and
frontal cortex, cingulate cortex, and striatum (Cardinal
Lewinsohn, 1973; Youngren and Lewinsohn, 1980).
et al, 2002). In the former study with healthy human
Additionally, it is notable that the reported effects were
participants (Park et al, 1994), tryptophan depletion also
apparent in an iterated, sequential PD game in which the
had a disproportionately greater impact on such associative
participants of the study made their choice about whether
learning on the first study day when participants completed
to cooperate or defect only after their playing partnerF
the task for the first time. The capacity to link (and
in fact, a computer playing a strict tit-for-tat strategyF
dynamically) relink stimuli to motivationally relevant
had indicated his/her response. In this case, cooperative
outcomes is critical for effective function within social
responses involved foregoing higher rewards available from
environments in which such linkages are apt to change
a defection response (ie 30p) in preference for smaller
rapidly and unpredictably (Rolls, 1996). Such conditions
rewards available from mutually cooperative responses (ie
clearly obtained on study day 1 on which participants were
20p). Behavioral analyses of iterated PD games have shown
required to assess the behavior of their partner and to
that a player has two incompatible objectives in these
adjust their own strategies in the light of varying outcomes
circumstances (Raichlin et al, 2001). On the one hand, he/
from each round of the PD game. These conditions were
she wishes to defect; on the other hand, he/she wishes to get
less apparent on day 2 when participants repeated the task
his/her partner to cooperate. However, their partner’s strict
with the same fictional (confederate) and real (computer)
tit-for-tat strategy ensures that any defection will incur a
retaliatory defection on the immediately following round of
These results shed light on the underlying role of
the game. At this point, the player will face a choice between
serotonin in mediating social cooperation in healthy human
becoming locked in an ongoing pattern of mutual defec-
participants, and highlight several directions for future
tions or be forced to cooperate (while his partner defects) in
research into this area, in addition to suggesting candidate
order to re-establish a pattern of mutual cooperation. In this
neuropsychological mechanisms for social dysfunction in
way, playing the iterated, sequential PD game, in which the
partner plays strict tit-for-tat, involves the exercise of self-
Brain-imaging studies have reported that mutual co-
control in forgoing the higher immediate reward produced
operative outcomes in the context of an iterated PD game of
by defecting when the partner has cooperated in favor of
a kind similar to that played in this study was associated
the immediately smaller (but cumulatively greater) rewards
with increased BOLD signal within the rostral anterior
associated with enduring mutual cooperation.
Effects of tryptophan depletion on PD game performanceRM Wood et al
Consistent with work indicating that tit-for-tat strategies
Bechara A, Tranel D, Damasio H, Damasio AR (1996). Failure to
cooperative behavior (Axelrod, 1984; Sheldon, 1999), the T
respond automatically to anticipated future outcomes following
+ treatment in the current study produced a clear
damage to prefrontal cortex. Cerebral Cortex 6: 215–225.
preference for cooperation on both the first (0.7370.06)
Biggio G, Fadda F, Fanni P, Tagliamonte A, Gessa G-L (1974).
and second day (0.6470.09), suggesting that performance
Rapid depletion of serum tryptophan, brain tryptophan,
was supported by a strong representation of the longer-term
serotonin and 5-hyrdoxyindoleacetic acid by a tryptophan-freediet. Life Sci 14: 1321–1329.
benefits of mutual cooperation. Serotonin has frequently
Bjork JM, Dougherty DM, Moeller FG, Swann AC (2000).
been associated with aspects of impulsivity in both clinical
Differential behavioral effects of plasma tryptophan depletion
(Soubrie, 1986; Virkkunen et al, 1994) and experimental
and loading in aggressive and nonaggressive men. Neuropsycho-
settings (Dalley et al, 2002), with demonstrations that
tryptophan depletion can impair impulse control in healthy
Bond A, Lader M (1974). The use of analogue scales in rating
controls (Walderhaug et al, 2002) and in individuals with a
subjective feelings. Br J Med Psychol 47: 211–218.
family history of alcoholism (Crean et al, 2002; LeMarquand
Boone C, de Brabander B, Carree M, de Jong G, van Olffen W, van
et al, 1999). Other data have suggested that depletion of
Witteloostuijn A (2002). Locus of control and learning to cooperate
central serotonin impairs the capacity to delay gratification
in a prisoner’s dilemma game. Person Indiv Differ 32: 929–946.
(Denk et al, 2005; Mobini et al, 2000; Wogar et al, 1993).
Brown GL, Goodwin FK, Ballenger JC, Goyer PF, Major LF (1979).
Aggression in humans correlates with cerebrospinal fluid amine
Consequently, the present findings suggest that tryptophan
metabolites. Psychiatry Res 1: 131–139.
depletion significantly weakens the influence of the longer-
Buss AP, Perry MP (1992). The aggression questionnaire. J Person
term rewards associated with mutual cooperative strategies,
perhaps represented within circuitry encompassing the
Cardinal R, Parkinson J, Hall J, Everitt B (2002). Emotion and
orbitofrontal cortex (Bechara et al, 1996); thereby, under-
motivation: the role of the amygdala, ventral striatum, and
mining the basis for sustained reciprocation. This hypo-
prefrontal cortex. Neurosci Biobehav Rev 26: 321–352.
thesis may provide a basis for understanding how
Carpenter LL, Anderson GM, Pelton GH, Gudin JA, Kirwin PDS,
impulsiveness in personality disorders, associated with
Price LH et al (1998). Tryptophan depletion during continuous
serotonergic dysfunction, undermines social function
CSF sampling in healthy human subjects. Neuropsychopharma-
through an inadequate representation of the future
Clarke HF, Dalley JW, Crofts HS, Robbins TW, Roberts AC (2004).
reward-value of reciprocal cooperation. It also predicts
Cognitive inflexibility after prefrontal serotonin depletion.
that serotonergic manipulations will be most potent in
determining social interactions, such as those involving
Cleare AJ, Bond AJ (1995). The effect of tryptophan depletion
tit-for-tat interactions, which pit immediate and delayed
and enhancement on subjective and behavioural aggression in
prospects for reinforcement against each other; by contrast,
normal male subjects. Psychopharmacology 118: 72–81.
serotonergic manipulations in the context of other PD
Coccaro EF, Kavoussi RJ (1997). Fluoxetine and impulsive
games involving other kinds of playing strategies may not
aggressive behavior in personality-disordered subjects. Arch
In summary, we have demonstrated that depriving
Coull JT, Middleton HC, Robbins TW, Sahakian BJ (1995).
healthy participants of l-tryptophan reduced cooperative
Contrasting effects of clonidine and diazepam on tests of workingmemory and planning. Psychopharmacology 120: 311–321.
behavior and the tendency of mutual cooperation to
Crean J, Richards JB, de Wit H (2002). Effect of tryptophan
promote further cooperation. These results provide a
depletion on impulsive behaviour in men with or without a
starting point for testing novel hypotheses concerning the
family history of alcoholism. Behav Brain Res 136: 349–357.
role of serotonin in social cooperation and its dysfunction
Dalley JW, Theobald DE, Eagle DM, Passetti F, Robbins TW (2002).
in psychiatric disorders. These include (i) a failure to find
Deficits in impulse control associated with tonically-elevated
cooperation with others adequately reinforcing and (ii)
serotonergic function in rat prefrontal cortex. Neuropsycho-
weakening the strength of long-term gains (associated with
mutual cooperation) over short-term immediate rewards
Denk F, Walton ME, Jennings KA, Sharp T, Rushworth MF,
(associated with defection following cooperative responses
Bannerman DM (2005). Differential involvement of serotonin
and dopamine systems in cost–benefit decisions about delay oreffort. Psychopharmacology 179: 587–596.
Dubini ABM, Polin V (1997). Do noradrenaline and serotonin
differentially affect social motivation and behaviour? European
Neuropsychopharmacology 7(Suppl 1): S49–S55.
This research was funded by an independent award from
Elliott R, Sahakian BJ, Matthews K, Bannerjea A, Rimmer J,
the Biotechnology and Biological Sciences Research Council
Robbins TW (1997). Effects of methylphenidate on spatialworking memory and planning in healthy young adults.
(BBSRC; UK) to Robert D Rogers (S20137).
First MB, Spitzer RL, Gibbon M, Williams JBW (2002). Structured
Clinical Interview for DSM-IV-TR Axis I Disorders, ResearchVersion, Patient Edition. (SCID-I/P). Biometrics Research: New
Aronson S, Black J, McDougle C, Be S, Jatlow P, Heninger G et al
(1995). Serotonergic mechanisms of cocaine effects in humans.
Fletcher PJ, Korth KM, Chambers JW (1999). Selective destruction
of brain serotonin neurons by 5,7-dihydroxytryptamine in-
Axelrod R (1984). The Evolution of Cooperation. Penguin Books:
creases responding for a conditioned reward. Psychopharma-
Axelrod R, Hamilton WD (1981). The evolution of cooperation.
Fossi L, Faravelli C, Paoli M (1984). The ethological approach to the
assessment of depressive disorders. J Nerv Ment Dis 172: 332–341.
Effects of tryptophan depletion on PD game performance
Fuller RW (1996). The influence of fluoxetine on aggressive
influenced behaviors in vervet monkeys (Cercopithecus aethiops
behavior. Neuropsychopharmacology 14: 77–81.
sabaeus). Psychopharmacology 72: 241–246.
Giovanni PA, Placidi MA, Oquendo MA, Malone KM, Huang YY,
Redgrave P, Horrell RI (1976). Potentiation of central reward by
Ellis SP et al (2001). Aggressivity, suicide attempts, and
localised perfusion of acetylcholine and 5-hydroxytryptamine.
depression: relationship to cerebrospinal fluid monoamine
metabolite levels. Biol Psychiatry 50: 783–791.
Rilling JK, Gutman DA, Zeh TR, Guiseppe P, Berns GS, Kilts CD
Harrison A, Markou A (2001). Serotonergic manipulations both
(2002). A neural basis for social cooperation. Neuron 35:
potentiate and reduce brain stimulation reward in rats:
involvement of serotonin-1A receptors. J Pharmacol Exp Ther
Rilling JK, Sanfey AG, Aronson JA, Nystrom LE, Cohen JD (2004).
The neural correlates of theory of mind within interpersonal
Higley JD, Mehlman PT, Poland RE, Taub DM, Higley SB, Suomi SJ
interactions. Neuroimage 22: 1694–1703.
et al (1992). Cerebrospinal fluid monoamine and adrenal
Robbins TW, Cador M, Taylor J, Everitt B (1989). Limbic-striatal
correlates of aggression in free-ranging rhesus monkeys. Arch
interactions in reward-related processes. Neurosci Biobehav Rev
Higley JD, Mehlman PT, Poland RE, Taub DM, Vickers J, Suomi SJ
Robbins TW, Everitt BJ (1996). Neurobehavioural mechanisms of
et al (1996). CSF testosterone and 5-HIAA correlate with different
reward and motivation. Curr Opin Neurobiol 6: 228–236.
types of aggressive behaviors. Biol Psychiatry 40: 1067–1082.
Rogers RD, Blackshaw AJ, Middleton HC, Matthews K, Deakin
Knutson B, Panksepp J (1996). Effects of fluoxetine on play
JFW, Sahakian BJ et al (1999). Tryptophan depletion impairs
dominance in juvenile rats. Aggress Behav 22: 297–307.
stimulus-reward learning while methylphenidate disrupts atten-
Knutson B, Wolkowitz OW, Cole SW, Chan T, Moore EA, Johnson RC
tional control in healthy young adults: implications for the
et al (1998). Selective alteration of personality and social behaviour
monoaminergic basis of impulsive behaviour. Psychopharma-
by serotonergic intervention. Am J Psychiatry 155: 373–379.
LeMarquand DG, Benkelfat C, Pihl RO, Palmour RM, Young SN
Rogers RD, Tunbridge EM, Bhagwagar Z, Drevets W, Sahakian BJ,
(1999). Behavioral disinhibition induced by tryptophan deple-
Carter CS (2003). Tryptophan depletion alters the decision-
tion in nonalcoholic young men with multigenerational family
making of healthy participants through altered processing of
histories of paternal alcoholism. Am J Psychiatry 156: 1771–1779.
reward cues. Neuropsychopharmacology 28: 153–162.
Libet J, Lewinsohn PM (1973). The concept of social skill with
Rolls ET (1996). The orbitofrontal cortex. Philos Trans Roy Soc
special references to the behaviour of depressed persons.
J Consult Clin Psychol 40: 304–312.
Sasaki-Adams DM, Kelley AE (2001). Serotonin–dopamine inter-
Marsh DM, Dougherty DM, Moeller FG, Swann AC, Spiga R (2002).
actions in the control of conditioned reinforcement and motor
Laboratory-measured aggressive behavior of women: acute
behavior. Neuropsychopharmacology 25: 440–452.
tryptophan depletion and augmentation. Neuropsychopharma-
Sheldon KM (1999). Learning the lessons of tit-for-tat: even
competitors can get the message. J Person Soc Psychol 77:
Mobini S, Chiang TJ, Ho MY, Bradshaw C, Szabadi E (2000).
Effects of central 5-hydroxytryptamine depletion on sensitivity
Soubrie P (1986). Reconciling the role of serotonin neurons in
to delayed and probabilistic reinforcement. Psychopharmacology
human and animal behaviour. Behav Brain Sci 9: 319–362.
Trivers R (1971). The Evolution of Reciprocal Altruism. Q Rev Biol
Moore P, Hans-Peter L, Seifritz E, Clark C, Bhatti T, Kelsoe J et al
(2000). Clinical and physiological consequences of rapid
Trivers R (1985). Social Evolution. Benjamin/Cummings: Menlo
tryptophan depletion. Neuropsychopharmacology 23: 601–622.
Moskowitz DS, Cote S (1995). Do interpersonal traits predict
Tse WS, Bond AJ (2002a). Serotonergic intervention affects both
affect? A comparison of three models. J Person Soc Psychol 69:
social dominance and affiliative behaviour. Psychopharmacology
Moskowitz DS, Pinard G, Zuroff DC, Annable L, Young SN (2001).
Tse WS, Bond AJ (2002b). Differences in serotonergic and
The effect of tryptophan on social interaction in everyday life: a
noradrenergic regulation of human social behaviours. Psycho-
placebo-controlled study. Neuropsychopharmacology 25: 277–289.
Nelson HE (1982). National Adult Reading Test (NART) Test
Virkkunen M, Rawlings R, Tokola R, Poland RE, Guidotti A,
Nemeroff C et al (1994). CSF biochemistries, glucose metabo-
Park SB, Coul JT, McShane RH, Young AH, Sahakian BJ, Robbins
lism, and diurnal activity rhythms in alcoholic, violent offenders,
TW et al (1994). Tryptophan depletion in normal participants
fire setters, and healthy participants. Arch Gen Psychiatry 51:
produces selective impairments in learning and memory.
Walderhaug E, Lunde H, Nordvik JE, Landro NI, Refsum H,
Pruitt D, Kimmel M (1977). Twenty years of experimental gaming:
Magnusson A (2002). Lowering of serotonin by rapid trypto-
critique, synthesis, and suggestions for the future. Annu Rev
phan depletion increases impulsiveness in normal individuals.
Raichlin H, Brown J, Baker F (2001). Reinforcement and punish-
Watson D, Clark LA, Tellegen A (1988). Development and
ment in the Prisoner’s Dilemma game. Psychol Learn Motiv 40:
validation of brief measures of positive and negative affect:
The PANAS scales. J Person Soc Psychol 54: 1063–1070.
Raleigh MJ, Brammer GL, McGuire MT, Yuwiler A (1985).
Weissman MM (2000). Social functioning and the treatment of
Dominant social status facilitates the behavioral effects of
depression. J Clin Psychiatry 61(Suppl 1): 33–38.
serotonergic agonists. Brain Res 348: 274–282.
Wogar MA, Bradshaw CM, Szabadi E (1993). Effect of lesion of the
Raleigh MJ, McGuire MT, Brammer GL, Pollack DB, Yuwiler A
ascending 5-hydroxytryptaminergic pathways on choice between
(1991). Serotonergic mechanisms promote dominance acquisi-
delayed reinforcers. Psychopharmacology 111: 239–243.
tion in adult male monkeys. Brain Res 559: 181–190.
Youngren MA, Lewinsohn PM (1980). The functional relation
Raleigh MJ, Yuwiler A, Brammer GL, McGuire MT, Geller E,
between depression and problematic interpersonal behavior.
Flannery JW (1981). Peripheral correlates of serotonergically-
NUVARING NAME OF THE MEDICINAL PRODUCT NuvaRing® 2. QUALITATIVE QUANTITATIVE COMPOSITION NuvaRing contains 11.7 mg etonogestrel and 2.7 mg ethinylestradiol. The ring releases etonogestrel and ethinylestradiol at an average amount of 0.120 mg and 0.015 mg, respectively per 24 hours, over a period of 3 weeks. For excipients, see 6.1. 3. PHARMACEUTICAL NuvaRing is flexible,
AMERICAN SOCIETY FOR REPRODUCTIVE MEDICINE Formerly The American Fertility Society 1209 MONTGOMERY HIGHWAY • BIRMINGHAM, ALABAMA 35216-2809 • TEL (205)978-5000 • FAX (205)978-5005 • E-MAIL [email protected] PATIENT’S FACT SHEET: PROLACTIN EXCESS PROLACTIN: A MILK-PRODUCING HORMONE Prolactin is a hormone secreted by the pituitary gland which is located at the base of the brain. I
 Neuropsychopharmacology (2006) 31, 1075–1084& 2006 Nature Publishing Group
Effects of Tryptophan Depletion on the Performance of anIterated Prisoner’s Dilemma Game in Healthy Adults
Richard M Wood1, James K Rilling2, Alan G Sanfey2, Zubin Bhagwagar3 and Robert D Rogers*,1
University Department of Psychiatry, University of Oxford, Oxford, UK; 2Departments of Anthropology, Psychiatry and Behavioral Sciences,
Emory University, Atlanta, GA, USA; 3Department of Psychiatry, Yale University, New Haven, CT, USA
Adaptive social behavior often necessitates choosing to cooperate with others for long-term gains at the expense of noncooperative
behaviors giving larger immediate gains. Although little is know about the neural substrates that support cooperative over
noncooperative behaviors, recent research has shown that mutually cooperative behavior in the context of a mixed-motive game, the
Prisoner’s Dilemma (PD), is associated with increased neural activity within reinforcement circuitry. Other research attests to a role for
serotonin in the modulation of social behavior and in reward processing. In this study, we used a within-subject, crossover, double-blind
design to investigate performance of an iterated, sequential PD game for monetary reward by healthy human adult participants following
ingestion of an amino-acid drink that either did (T + ) or did not (TÀ) contain l-tryptophan. Tryptophan depletion produced significant
reductions in the level of cooperation shown by participants when playing the game on the first, but not the second, study days. This
effect was accompanied by a significantly diminished probability of cooperative responding given previous mutually cooperative behavior.
Neuropsychopharmacology (2006) 31, 1075–1084& 2006 Nature Publishing Group
Effects of Tryptophan Depletion on the Performance of anIterated Prisoner’s Dilemma Game in Healthy Adults
Richard M Wood1, James K Rilling2, Alan G Sanfey2, Zubin Bhagwagar3 and Robert D Rogers*,1
University Department of Psychiatry, University of Oxford, Oxford, UK; 2Departments of Anthropology, Psychiatry and Behavioral Sciences,
Emory University, Atlanta, GA, USA; 3Department of Psychiatry, Yale University, New Haven, CT, USA
Adaptive social behavior often necessitates choosing to cooperate with others for long-term gains at the expense of noncooperative
behaviors giving larger immediate gains. Although little is know about the neural substrates that support cooperative over
noncooperative behaviors, recent research has shown that mutually cooperative behavior in the context of a mixed-motive game, the
Prisoner’s Dilemma (PD), is associated with increased neural activity within reinforcement circuitry. Other research attests to a role for
serotonin in the modulation of social behavior and in reward processing. In this study, we used a within-subject, crossover, double-blind
design to investigate performance of an iterated, sequential PD game for monetary reward by healthy human adult participants following
ingestion of an amino-acid drink that either did (T + ) or did not (TÀ) contain l-tryptophan. Tryptophan depletion produced significant
reductions in the level of cooperation shown by participants when playing the game on the first, but not the second, study days. This
effect was accompanied by a significantly diminished probability of cooperative responding given previous mutually cooperative behavior. Effects of tryptophan depletion on PD game performance
in reward and the processing of motivationally significant
Chronic citalopram was associated with increased choices
information (Robbins and Everitt, 1996), suggesting that
of the more generous pay-offs and increased affiliative
socially cooperative behavior is supported by activity within
messages sent to the playing partner (Tse and Bond, 2002a).
Effects of tryptophan depletion on PD game performance
in reward and the processing of motivationally significant
Chronic citalopram was associated with increased choices
information (Robbins and Everitt, 1996), suggesting that
of the more generous pay-offs and increased affiliative
socially cooperative behavior is supported by activity within
messages sent to the playing partner (Tse and Bond, 2002a).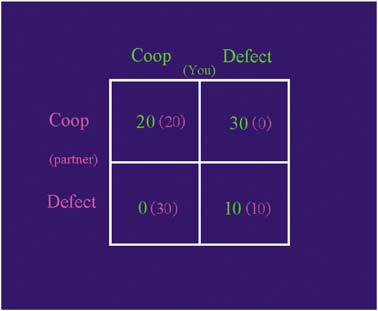
 Effects of tryptophan depletion on PD game performanceRM Wood et al
game, each participant was introduced to a gender-matched‘partner’Fin reality, an experimenter-confederate. It was
The study consisted of a within-subject, crossover, double-
explained to the participant that his/her computer was
blind design. Each participant ingested an amino-acid drink
networked to a computer in another room in which their
on 2 study days, separated by not less than 1 week. One
partner would play. The experimenter left to take the
drink contained l-tryptophan (T + ), the other drink did not
confederate to their room, returning a few minutes later to
(TÀ). The order of these treatments was counterbalanced
talk the participant through the PD game instructions.
Effects of tryptophan depletion on PD game performanceRM Wood et al
game, each participant was introduced to a gender-matched‘partner’Fin reality, an experimenter-confederate. It was
The study consisted of a within-subject, crossover, double-
explained to the participant that his/her computer was
blind design. Each participant ingested an amino-acid drink
networked to a computer in another room in which their
on 2 study days, separated by not less than 1 week. One
partner would play. The experimenter left to take the
drink contained l-tryptophan (T + ), the other drink did not
confederate to their room, returning a few minutes later to
(TÀ). The order of these treatments was counterbalanced
talk the participant through the PD game instructions. Effects of tryptophan depletion on PD game performance
Table 1 Monetary Payoffs for Four Different Strategies Over the
tion of the data from the PD games.) Where the effects of
Course of the Iterated, Sequential Prisoner’s Dilemma (PD) Game
the TÀ and T + treatments were different on the first and
second study days, as indicated by significant two-wayinteractions between treatment and treatment order,
dependent measures were subject to supplementary ANO-VAs for each study day separately (to investigate simple
Both you and your partner cooperate each round
interaction effects), with gender and treatment (T + vs TÀ)
Your partner cooperates and you defect each round
as two between-subject factors and game as a single within-
Your partner defects and you cooperate each round
Both you and your partner defect each round
‘You’ ¼ participant; ‘Partner’ ¼ computer program playing tit-for-tat.
Effects of tryptophan depletion on PD game performance
Table 1 Monetary Payoffs for Four Different Strategies Over the
tion of the data from the PD games.) Where the effects of
Course of the Iterated, Sequential Prisoner’s Dilemma (PD) Game
the TÀ and T + treatments were different on the first and
second study days, as indicated by significant two-wayinteractions between treatment and treatment order,
dependent measures were subject to supplementary ANO-VAs for each study day separately (to investigate simple
Both you and your partner cooperate each round
interaction effects), with gender and treatment (T + vs TÀ)
Your partner cooperates and you defect each round
as two between-subject factors and game as a single within-
Your partner defects and you cooperate each round
Both you and your partner defect each round
‘You’ ¼ participant; ‘Partner’ ¼ computer program playing tit-for-tat.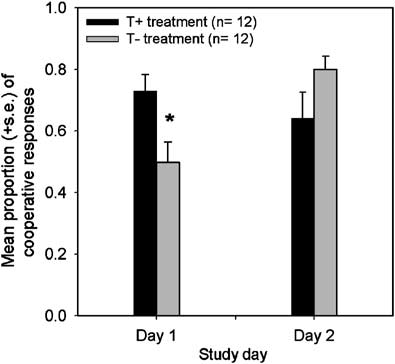
 Effects of tryptophan depletion on PD game performanceRM Wood et al
TÀ treatment on day 2 (0.7370.05 to 0.8070.04; t (two-
However, analysis of each study day separately showed that
tailed) ¼ À0.98, df ¼ 11, po0.35).
Effects of tryptophan depletion on PD game performanceRM Wood et al
TÀ treatment on day 2 (0.7370.05 to 0.8070.04; t (two-
However, analysis of each study day separately showed that
tailed) ¼ À0.98, df ¼ 11, po0.35). Effects of tryptophan depletion on PD game performance
treatment (13.5471.02 vs 12.6370.69) (Fso1.00). There
TÀ compared to the T + treatment on either study day (see
were no significant differences in the state positive or
Table 2). Moreover, the reduced cooperation of those
negative affect of the T + and TÀ participants at baseline
participants tested after the TÀ treatment on study day 1
or + 5 h on either study day 1 or day 2 (all Fso1.00;
cannot be attributed to an increase in state aggression in
see Table 2). Entering state positive and negative affect
comparison with those tested after the T + treatment. These
at + 5 h on day 1 as covariates did not abolish the
two groups of participants were matched on the multiple
significant reduction in cooperative responses associated
facets of aggression measured by the VAS (Bond and Lader,
with the TÀ compared to T + treatments (F(1, 18) ¼ 6.70,
1974) at baseline, and showed broadly similar changes
during the interval between baseline and playing the PDgame + 5 h later. At that time, 11 of the 13 VAS showedlower scores associated with the TÀ treatment compared to
the T + treatment and only two showed higher scores (see
Overall, self-reported annoyance was significantly decreased
Table 2). Indeed, the single significant treatment difference
after the TÀ compared to the T + treatment (21.7772.62 vs
(uncorrected for multiple comparisons) at the time of
26.0473.49) (F(1, 20) ¼ 4.88, po0.05); self-reported disgust
playing the PD game on day 1 indicated that participants
was also reduced after the TÀ treatment compared to the
tested after the TÀ treatment rated themselves as signifi-
T + treatment (21.8172.74 vs 29.1073.64) (F(1, 20) ¼ 5.48,
cantly less rebellious than those tested after the T +
po0.05). Comparisons of ratings at + 5 h on study day 1,
treatment. In summary, there is no evidence to suggest
taken just before playing the PD game, revealed that
that the reduced cooperation associated with the TÀ treat-
rebelliousness was significantly lower following the TÀ
ment on the first study day is explicable by differences in
state affect, or increased state aggressivity, consequent
state furiousness showed a similar trend (F(1, 20) ¼ 3.48,
to tryptophan depletion. Rather, the data indicate that
p ¼ 0.08). There were no other significant treatment-related
serotonin can affect social cooperative behavior indepen-
differences in state aggression ratings at baseline or + 5 h
dently of the gross changes in emotional state that might be
detectable by self-report measures of positive or negativeaffect including state aggression.
Effects of tryptophan depletion on PD game performance
treatment (13.5471.02 vs 12.6370.69) (Fso1.00). There
TÀ compared to the T + treatment on either study day (see
were no significant differences in the state positive or
Table 2). Moreover, the reduced cooperation of those
negative affect of the T + and TÀ participants at baseline
participants tested after the TÀ treatment on study day 1
or + 5 h on either study day 1 or day 2 (all Fso1.00;
cannot be attributed to an increase in state aggression in
see Table 2). Entering state positive and negative affect
comparison with those tested after the T + treatment. These
at + 5 h on day 1 as covariates did not abolish the
two groups of participants were matched on the multiple
significant reduction in cooperative responses associated
facets of aggression measured by the VAS (Bond and Lader,
with the TÀ compared to T + treatments (F(1, 18) ¼ 6.70,
1974) at baseline, and showed broadly similar changes
during the interval between baseline and playing the PDgame + 5 h later. At that time, 11 of the 13 VAS showedlower scores associated with the TÀ treatment compared to
the T + treatment and only two showed higher scores (see
Overall, self-reported annoyance was significantly decreased
Table 2). Indeed, the single significant treatment difference
after the TÀ compared to the T + treatment (21.7772.62 vs
(uncorrected for multiple comparisons) at the time of
26.0473.49) (F(1, 20) ¼ 4.88, po0.05); self-reported disgust
playing the PD game on day 1 indicated that participants
was also reduced after the TÀ treatment compared to the
tested after the TÀ treatment rated themselves as signifi-
T + treatment (21.8172.74 vs 29.1073.64) (F(1, 20) ¼ 5.48,
cantly less rebellious than those tested after the T +
po0.05). Comparisons of ratings at + 5 h on study day 1,
treatment. In summary, there is no evidence to suggest
taken just before playing the PD game, revealed that
that the reduced cooperation associated with the TÀ treat-
rebelliousness was significantly lower following the TÀ
ment on the first study day is explicable by differences in
state affect, or increased state aggressivity, consequent
state furiousness showed a similar trend (F(1, 20) ¼ 3.48,
to tryptophan depletion. Rather, the data indicate that
p ¼ 0.08). There were no other significant treatment-related
serotonin can affect social cooperative behavior indepen-
differences in state aggression ratings at baseline or + 5 h
dently of the gross changes in emotional state that might be
detectable by self-report measures of positive or negativeaffect including state aggression. Effects of tryptophan depletion on PD game performanceRM Wood et al
Table 2 Subjective Effects for Participants Who Drank an Amino-Acid Drink Containing l-Tryptophan (T+) and Participants Who Drankan Amino-Acid Drink not Containing l-Tryptophan (TÀ) on Study Day 1 and Day 2
State positive and negative affect (PANAS). Visual analog scales for state aggression (VAS). * ¼ F(1,20) ¼ 5.83, po0.05.
Effects of tryptophan depletion on PD game performanceRM Wood et al
Table 2 Subjective Effects for Participants Who Drank an Amino-Acid Drink Containing l-Tryptophan (T+) and Participants Who Drankan Amino-Acid Drink not Containing l-Tryptophan (TÀ) on Study Day 1 and Day 2
State positive and negative affect (PANAS). Visual analog scales for state aggression (VAS). * ¼ F(1,20) ¼ 5.83, po0.05. Effects of tryptophan depletion on PD game performance
some involving tryptophan depletion (Park et al, 1994).
Effects of tryptophan depletion on PD game performance
some involving tryptophan depletion (Park et al, 1994). Effects of tryptophan depletion on PD game performanceRM Wood et al
Consistent with work indicating that tit-for-tat strategies
Bechara A, Tranel D, Damasio H, Damasio AR (1996). Failure to
cooperative behavior (Axelrod, 1984; Sheldon, 1999), the T
respond automatically to anticipated future outcomes following
+ treatment in the current study produced a clear
damage to prefrontal cortex. Cerebral Cortex 6: 215–225.
Effects of tryptophan depletion on PD game performanceRM Wood et al
Consistent with work indicating that tit-for-tat strategies
Bechara A, Tranel D, Damasio H, Damasio AR (1996). Failure to
cooperative behavior (Axelrod, 1984; Sheldon, 1999), the T
respond automatically to anticipated future outcomes following
+ treatment in the current study produced a clear
damage to prefrontal cortex. Cerebral Cortex 6: 215–225. Effects of tryptophan depletion on PD game performance
Fuller RW (1996). The influence of fluoxetine on aggressive
influenced behaviors in vervet monkeys (Cercopithecus aethiops
behavior. Neuropsychopharmacology 14: 77–81.
Effects of tryptophan depletion on PD game performance
Fuller RW (1996). The influence of fluoxetine on aggressive
influenced behaviors in vervet monkeys (Cercopithecus aethiops
behavior. Neuropsychopharmacology 14: 77–81.