Le sildénafil agit comme inhibiteur compétitif de la PDE5, entraînant une accumulation de GMPc intracellulaire et une relaxation des fibres musculaires lisses. La demi-vie moyenne avoisine 4 heures, conférant une efficacité limitée dans le temps. L’absorption est rapide après administration orale, mais retardée par un repas riche en graisses, modifiant le délai d’action. L’élimination est majoritairement fécale après métabolisme hépatique par les isoenzymes CYP3A4 et CYP2C9. Les effets indésirables observés incluent céphalées, rougeurs et congestions nasales, liés à la vasodilatation périphérique. Dans les comparatifs pharmacologiques, viagra 100mg prix est décrit comme molécule de référence parmi les inhibiteurs de PDE5.
Genetic calcium detector
Commentary Cell Science Reviews Vol 5 No 2 ISSN 1742-8130 Single spike detection with a genetic calcium sensor Mika Mizunuma 1, Megumi Seki 1, Atsushi Usami 1 & Yuji Ikegaya 1,2
1 Laboratory of Chemical Pharmacology, Graduate School of Pharmaceutical Sciences, The University of Tokyo, 7-3-1 Hongo,
2 Precursory Research for Embryonic Science and Technology (PRESTO), Japan Science and Technology Agency, 5 Sanbancho
Received 15th October Cell Science 2008
Although genetically encoded calcium probes have started to be used in the live recording of neuronal activity, their low sensitivity and expression levels have not allowed the detection of single action potentials. Recently, Wallace et al. overcame this considerable problem by delivering the sensor protein D3cpv using an adeno-associated viral vector.
The brain exerts its high-order function through the dynamic interactions of myriad neurons. Individual neurons work as functional elements within a topologically defined network. The mechanisms underlying the neural information processing, however, have not been fully elucidated, principally due to the experimental difficulty in monitoring large neuronal populations at the single cell resolution for long periods. To solve this problem, fluorescent calcium indicator proteins (FCIPs), i.e., genetically encoded reporters which alter their fluorescent properties in response to an increase in calcium ion activities, have recently been employed [1]. In neurons where the FCIP gene has been introduced under the downstream influence of an appropriate promoter, spike activity may be captured as a change in fluorescence intensity, as action potentials evoke transient calcium elevations through an activation of voltage-sensitive calcium channels. In a recent report [2], an experimental group has fine-tuned the properties of FCIP and enhanced its expression level in order to reliably detect single spikes within both in vitro and in vivo preparations.
FCIPs have significant advantages over chemically synthesized fluorescent indicators such as fura-2, fluo-4, or Oregon green. First, once sensor genes are introduced into neurons, for example in transgenic animals, the stable expression of the sensor proteins allows long-term recording from individual neurons, whereas with chemical dyes it is impractical to monitor the activity of the same neurons over days or weeks. More critically, widely used chemical indicators tend to decrease their loading efficiency in mature neural tissues, and hence experiments employing adult animal brains prove virtually impossible to record activity from in many cases. Another advantage of FCIPs is the ability to attain specific expression within certain types of cells or tissues under the control of appropriate promoters. Indeed, in a recent report TN-XXL, a novel FCIP, was introduced into neurons within the mouse visual cortex, and the same identical neurons were repetitively recorded from for a couple of weeks [3].
FCIPs are classified into two types, based on their structure and principles of operation, namely (i) single fluorescent protein-based sensors, and (ii) fluorescence resonance energy transfer (FRET)-based sensors [1]. Both types of FCIPs employ green fluorescent protein (GFP) or its derivatives to serve as a fluorescence emitter and use a core peptide of calcium-binding proteins, including calmodulin (CaM) and Toroponin C, to function as calcium detectors (Figure 1).
The first FCIP group (i) includes a set of G-CaMPs that absorb blue light and emit green fluorescence. In G-CaMPs, a circular permutated enhanced GFP (cpEGFP) molecule is linked to CaM and the M13 fragment of myosin light chain kinase. Calcium binding to the CaM moiety triggers the interaction between CaM and M13 domains and thereby changes the three-dimensional conformation of cpEGFP, resulting in a ~5-fold increase in fluorescence. In G-CaMP1.6, the brightness is augmented by point mutations within G-CaMP [4], and the stability at body temperature is enhanced in G-CaMP2 [5, 6].
Figure 1. Schematic architecture of fluorescent calcium indicator proteins (FCIPs).A. The structure of G-CaMP, which consists of circularly permutated enhanced GFP (cpEGFP), CaM, and its target peptide M13. B. (1) The structure of yellow chameleon, a FRET-based sensor. In
the absence of calcium ions, the donor CFP mainly generates fluorescence, whereas in the presence of calcium ions, the binding of CaM to M13 augments the FRET efficiency between CFP and YFP, and the acceptor YFP increases in fluorescence. (2) Structure of D3cpV. The FRET efficiency and the signal-to-noise ratio are both improved, as compared to yellow cameleon.
The second FCIP group (ii) includes cameleon and yellow cameleon. Genetic modifications of GFP produced a variety of useful fluorescent mutants, such as blue fluorescent protein (BFP), cyan fluorescent protein (CFP), and yellow fluorescent protein (YFP). Cameleon is the first FRET-based protein sensor, binding BFP and GFP via a linker of CaM and M13 [7]. Following cameleon, yellow cameleon was developed by replacing chameleon’s BFP and GFP with CFP and YFP, respectively [8]. Binding to calcium ions changes the direction and distance between two fluorescent groups, leading to an increase of the FRET efficiency. The increased energy transition from the excited donor CFP to the acceptor YFP results in a rise of the fluorescence intensity. A core advantage of FRET is the capacity for ratiometric imaging at different wavelengths, which enables relatively stable and highly sensitive calcium detection even within gradually photobleached or shape-changing samples. Various combinations of donor and acceptor proteins have been designed to produce different levels of fluorescence intensity. Calcium sensor parts have also been improved to attain higher stability against changes in temperature or pH, and to reduce undesirable interactions with other endogenous molecules.
The sensor D3cpv is one of these improved FRET-based sensors. In a previous study, the precise locations of several point mutations in M13 were computationally determined to suppress its interaction with endogenous CaM [9]. Among these candidates, the so-called design III was adopted in D3cpv. To obtain FRET, two tails of D3cpv were fused to enhanced CFP and circularly permutated Venus. Due to its relatively high affinity to calcium ions, D3cpv is responsive to a relatively small change in the concentration of free calcium, such that it detects spontaneous spike activity within hippocampal neurons in vitro, but was still however unable to detect single spikes, due to a lack of sensitivity.
In order to enable the detection of single spikes, the authors tried to increase the expression level of this sensor protein. They used recombinant adeno-associated virus (rAAVs) to introduce the D3cpV gene. The rAAV system is known to bring about high protein expression levels with little resulting damage to the infected cells [10]. The authors used the rAAV gene delivery technique and succeeded in detecting single spike activity with high fidelity in both hippocampal pyramidal cells in vitro and somatosensory cortex in vivo. They also combined this technique with the tetracycline promoter system to further enhance the expression level. This
gene expression system seems to work well, especially in some in vitro experiments.
However, at present the FRET kinetics of D3cpv are not fast enough to separate individual spikes at firing frequencies greater than 1 Hz. Thus, the sensor needs to be further improved so as to display more rapid, temporally defined responses to the calcium fluctuations which arise from individual action potentials. Nonetheless, this new optical system, which offers single spike sensitivity, is already proving a promising tool for the recording of network activity in large populations of defined neurons over long time intervals at a cellular level and fine temporal resolution.
References
1. Miyawaki A (2005) Innovations in the imaging of brain functions using fluorescent proteins. Neuron 48: 189-199.
2. Wallace DJ, Zum Alten Borgloh SM, Astori S, Yang Y, Bausen M, Kügler S, Palmer AE, Tsien RY, Sprengel R, Kerr JN, Denk W, Hasan MT (2008) Single-spike detection in vitro and in vivo with a genetic calcium sensor. Nat Methods 5:797-804.
3. Mank M, Santos AF, Direnberger S, Mrsic-Flogel TD, Hofer SB, Stein V, Hendel T, Reiff DF, Levelt C, Borst A, Bonhoeffer T, Hübener M, Griesbeck O (2008) A genetically encoded calcium indicator for chronic in vivo two-photon imaging. Nature Methods 5:805-811.
4. Ohkura M, Matsuzaki M, Kasai H, Imoto K, Nakai J (2005) Geneticallyencoded bright Ca2+ probe applicable for dynamic Ca2+ imaging of dendritic spines. Anal Chem 77:5861-5869.
5. Díez-García J, Matsushita S, Mutoh H, Nakai J, Ohkura M, Yokoyama J, Dimitrov D, Knöpfel T (2005) Activation of cerebellar parallel fibers monitored in transgenic mice expressing a fluorescent Ca2+ indicator protein. Eur J Neurosci 22:627-635.
6. Tallini YN, Ohkura M, Choi BR, Ji G, Imoto K, Doran R, Lee J, Plan P, Wilson J, Xin HB, Sanbe A, Gulick J, Mathai J, Robbins J, Salama G, Nakai J, Kotlikoff MI (2006) Imaging cellular signals in the heart in vivo: cardiac expression of the high-signal Ca2+ indicator GCaMP2. Proc Natl Acad Sci U S A 103:4753-4758.
7. Miyawaki A, Llopis J, Heim R, McCaffery JM, Adams JA, Ikura M, Tsien RY (1997) Fluorescent indicators for Ca2+ based on green fluorescent proteins and calmodulin. Nature 388:882-887.
8. Nagai T, Yamada S, Tominaga T, Ichikawa M, Miyawaki A (2004) Expanded dynamic range of fluorescent indicators for Ca2+ by circularly permuted yellow fluorescent proteins. Proc. Natl. Acad. Sci. USA 101: 10554-10559
9. Palmer AE, Glacomello M, Kortemme T, Hires SA, Lev-Ram V, Baker David, Tsien RY (2006) Ca2+ indicators Based on Computationally Redesigned Calmodulin-Peptide Pairs. Chemistry & Biology 13:521-530.
10. Schnepp BC, Jensen RL, Chen CL, Johnson PR Clark KR (2005) Characterization of adeno-associated virus genomes isolated from human tissues. J. Virol 79:4793–14803.
Bewertung der Sartane im Vergleich zu ACE-Hemmstoffen Prof. Dr. med. Wolfgang Motz Klinikum Karlsburg Herz- und Diabeteszentrum Mecklenburg-Vorpommern Greifswalder Str. 11 17495 Karlsburg In den letzten 20 Jahren konnte gezeigt werden, dass das Enzym Angiotensin II eine Schlüsselrol e bei der arteriel en Hypertonie und der Herzinsuffizienz spielt. Eine Hemmung der Konversion von Angiotensin I
(under the guidance of Mayor Bobot Umali) Surgical team: National Committee on Sight Preservation headed Talipanan, Puerto Galera Aninuan, Puerto Galera Balatero, Puerto Galera Poblacion, Puerto Galera Palangan, Puerto Galera Paraway, San Teodoro Saclag, San Teodoro Sipit Saburan, San Teodoro Ambang, San Teodoro Lantuyan, Baco San Lorenzo, Baco Katarata, Baco Paraiso, Baco Baras, Baco Matahimik










 Commentary
Commentary 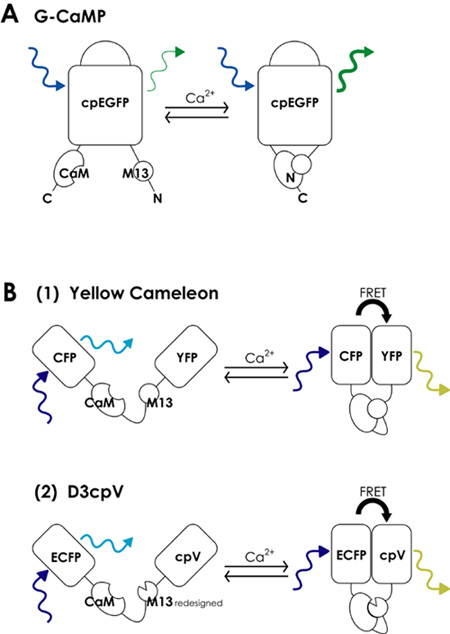 Figure 1. Schematic architecture of fluorescent calcium indicator proteins (FCIPs). A. The
Figure 1. Schematic architecture of fluorescent calcium indicator proteins (FCIPs). A. The