Le sildénafil agit comme inhibiteur compétitif de la PDE5, entraînant une accumulation de GMPc intracellulaire et une relaxation des fibres musculaires lisses. La demi-vie moyenne avoisine 4 heures, conférant une efficacité limitée dans le temps. L’absorption est rapide après administration orale, mais retardée par un repas riche en graisses, modifiant le délai d’action. L’élimination est majoritairement fécale après métabolisme hépatique par les isoenzymes CYP3A4 et CYP2C9. Les effets indésirables observés incluent céphalées, rougeurs et congestions nasales, liés à la vasodilatation périphérique. Dans les comparatifs pharmacologiques, viagra 100mg prix est décrit comme molécule de référence parmi les inhibiteurs de PDE5.
Saudecerebral.com.br
Effect of fish and fish oil-derived omega-3 fatty acids on lipid oxidation School of Medicine and Pharmacology, The University of Western Australia; Cardiovascular Research Centre; and The Western Australian Institute for Medical Research, Perth, Western Australia, Australia
There is evidence that omega-3 (ω3) fatty acids derived from fish and fish oils reduce the risk ofcardiovascular disease via mechanisms underlying atherosclerosis, thrombosis and inflammation. Despite these benefits, there has been concern that these fatty acids may increase lipid peroxidation. However, the in vivo data to date are inconclusive, due in part to limitations in the methodologies. Inthis regard, our findings using the measurement of F -isoprostanes, a reliable measure of in vivo
lipid peroxidation and oxidant stress, do not support adverse effects of ω3 fatty acids on lipidperoxidation. Keywords: Fish oil, omega-3 fatty acids, lipid oxidation, F -isoprostanes
There is now considerable evidence that a diet rich in ω3
dation and oxidative stress in the pathogenesis of cardio-
fatty acids derived from fish and fish oils, specifically
vascular disease.8 It is thought that oxidative stress and
eicosapentaenoic acid (EPA, 20:5 ω3) and docosa-
oxidized lipids play a critical part in the genesis and pro-
hexaenoic acid (DHA, 22:6 ω3), protects against athero-
gression of the atherosclerotic lesion.8 The hypothesis that
sclerotic heart disease, myocardial infarction and sudden
increased intake of ω3 fatty acids may lead to increased
death.1 ω3-Fatty acids have a wide range of biological
lipid peroxidation is based on the premise that fatty acid
effects, including benefits on lipoprotein metabolism,
oxidizability increases with an increase in the number of
platelet and endothelial function, blood pressure, vascu-
double bonds in the fatty acid chain.9 Whilst this may be
lar reactivity, cytokine production, coagulation and fibri-
true of in vitro studies of lipid peroxidation in homoge-
nolysis.1–3 Recent evidence also has demonstrated that,
neous solutions, in vivo systems are more complicated and
in humans, EPA and DHA have differential effects on
influenced by additional factors. In support of this, Visioli
lipids and lipoproteins,4 blood pressure5 and heart rate,5
et al.10 showed that, under equal conditions of oxidative
Published by Maney Publishing (c) W. S. Maney & Son Limited
stress, fatty acids oxidized at different rates and generated
Despite the benefits associated with increased ω3 fatty
different oxidation products, in a manner that was unre-
acid consumption, there remains a theoretical concern that
lated to their degree of unsaturation. Lipid peroxide levels
these fatty acids may increase the unsaturation index, con-
were in fact highest following oxidation of linoleic acid
sequent to the incorporation of EPA and DHA into mem-
(18:2 ω6).10 Arachidonic acid (20:4 ω6) and DHA gener-
branes and lipoproteins, leading to increased lipid
ated lower levels of lipid peroxides, with lowest levels
peroxidation.7 The significance of this relates to the fact
arising from oxidation of EPA. Formation of conjugated
that there is much evidence supporting a role for lipid oxi-
dienes was also maximal for 18:2 ω6. In particular, theproduction of conjugated dienes from 20:4 ω6 and EPAwas approximately 25% of that of 18:2 ω6. DHA oxida-
tion yielded only 10% of the conjugated dienes relative to
Correspondence to: Dr Trevor A. Mori, School of Medicine and
The data relating to the effects of ω3 fatty acids on lipid
Pharmacology, Medical Research Foundation Building, Box X 2213
peroxidation and oxidative stress in vivo are contradictory.7
GPO, Perth, Western Australia 6847, AustraliaTel: +61 8 9224 0273; Fax: +61 8 9224 0246;
These inconsistencies may be related to differences in the
populations studied, the quantity and presentation of the
gas chromatography mass spectrometry. In addition,plasma TBARS were significantly elevated following
ω3 fatty acids, whereas MDA was reduced. Whenplasma MDA levels were normalized to plasma polyun-saturated fatty acid concentrations, significant differ-ences were eliminated.25
Other measures of oxidative stress include electron
spin resonance detecting free radical species,24 measure-ment of antibodies to oxidized LDL31 and breath excre-tion of ethane and pentane.24 The latter assay, however,has yielded variable results in animals32 and humans27supplemented with ω3 fatty acids. Other literaturereports have shown that ω3 fatty acids had no adverseeffects on plasma protein oxidation26 and rendered ery-throcytes more resistant to haemolysis following oxida-tive challenge.33
On the basis of studies reporting adverse effects on
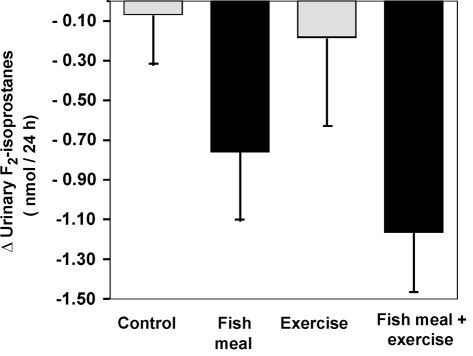
Fig. 1. Change in urinary F -isoprostane excretion from baseline to post-
intervention by intervention group. Mean ± SEM. P < 0.013 for the main
lipid peroxidation, some researchers have suggested that
effect of fish after adjustment for baseline values, using general linear
ω3 fatty acids may affect the antioxidant status and,
models (GLM). Reproduced from Mori et al.41 with permission.
therefore, should be taken in conjunction with vitaminsupplementation. The data from such studies, however,are inconclusive.16,27,34–37
ω3 fatty acids (fish versus encapsulated fish oils), the
Although most of the above-mentioned methods rep-
duration of the study, the study design, the antioxidant
resent different aspects of lipid oxidation and collec-
content of the supplement, or the composition of the
tively provide some knowledge of oxidative damage,
background diet. It also has also been suggested that the
none is considered a reliable measure of oxidative stress.
total concentration of polyunsaturates, rather than the
In this regard, the F -isoprostanes are prostaglandin-like
unsaturation index, may be more important in determin-
metabolites of free radical peroxidation of arachidonic
ing lipid peroxidation.11 However, the most plausible
acid38 and there is now good evidence that they provide a
explanation for the inconsistency between studies is dif-
reliable measure of in vivo oxidative stress.39,40 In sup-
ferences in the methodologies employed to assess lipid
port of this, elevated F -isoprostanes have been reported
in animal models of free radical injury, in human condi-
Much of the literature regarding the effect of ω3 fatty
tions associated with increased oxidative stress, and in
acids on lipid peroxidation is based on indirect and/or
non-specific assays. Studies utilizing the oxidizability of
Using F -isoprostanes, measured by gas chromatogra-
LDL have shown either enhanced,12–17 or reduced, or no
phy–mass spectrometry, we have demonstrated that
effect,11,18–22 of ω3 fatty acids. In this assay, LDL is iso-
these metabolites are significantly reduced following
lated from plasma and then subjected to oxidative condi-
consumption of ω3 fatty acids taken as fish oils or in fish
tions. It is feasible that in patients on various
meals. We showed that fish meals providing approxi-
Published by Maney Publishing (c) W. S. Maney & Son Limited
medications, LDL oxidizability could be affected by par-
mately 3.6 g/day of ω3 fatty acids for 8 weeks to Type 2
tition of drugs into the LDL. Indeed, a divergence has
diabetic patients, significantly (P = 0.013) reduced uri-
been shown between the measurement of LDL oxidative
nary F -isoprostanes by 20% (Fig. 1).41 Relative to a
susceptibility and urinary F -isoprostanes as a measure
control group, urinary F -isoprostanes were reduced by
0.83 nmol/24-h. This effect was independent of age,
One of the most common methods for assessment of
gender, body weight change and the increase in ω3 fatty
lipid peroxidation measures thiobarbituric acid reactive
acids or the fall in ω6 fatty acids in plasma, platelets and
substances (TBARS) by a colorimetric assay. This assay
has been widely criticized on the basis of its lack of
We recently have shown that cord plasma and urinary
specificity and results need to be interpreted with care.24
F -isoprostanes were reduced in infants whose mother
Studies of ω3 fatty acids using this methodology have
received fish oil during pregnancy.42 Pregnant atopic
shown either elevated levels16,17,25–28 or no change29,30 in
women received 4 g daily fish oil or olive oil from 20
TBARS. The limitation of this assay was exemplified by
weeks’ gestation. Cord plasma F -isoprostanes were sig-
Higdon et al.25 who showed that, in post-menopausal
nificantly lower (P < 0.001) in the offspring of women
women given ω3 fatty acids, plasma TBARS were 10-
who had taken fish oil during pregnancy compared with
fold higher than malondialdehyde (MDA) measured by
those who took olive oil (Fig. 2). These differences were
Effect of fish and fish oil-derived omega-3 fatty acids on lipid oxidation
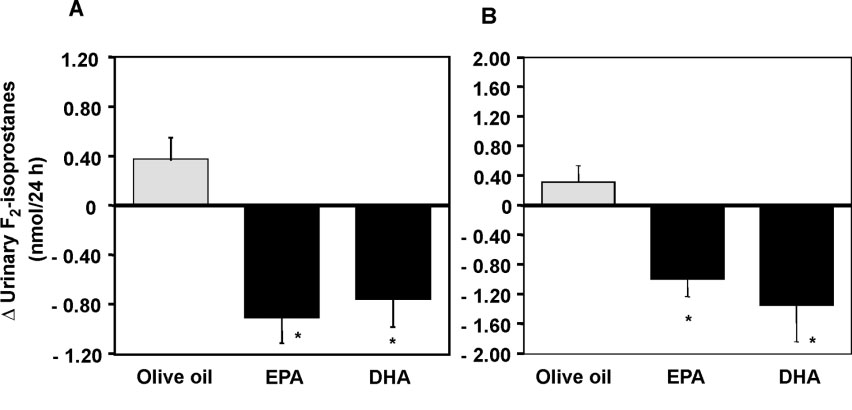
mildly hyperlipidaemic men, supplementation with 4 gdaily of purified EPA or DHA for 6 weeks decreased post-intervention urinary F -isoprostane levels by 27% follow-
ing EPA (1.24 nmol/24-h, P < 0.0001) and 26% followingDHA (1.20 pmol/24-h, P < 0.0001), relative to an olive oilcontrol group, after adjusting for baseline values (Fig. 3A).44 In a study of similar design in treated hypertensiveType 2 diabetic patients, we showed that post-interventionurinary F -isoprostanes were reduced 19% by EPA (P =
0.017) and 20% by DHA (P = 0.014), relative to an oliveoil group (Fig. 3B).45
In each of these studies,41,42,44,45 the changes in F -iso-
prostanes were unrelated to changes in EPA, DHA, 20:4
ω6, total ω3 or ω6 fatty acids. This lack of associationwith changes in fatty acids is noteworthy, in view of the
Fig. 2. Cord plasma F -isoprostane excretion in neonates whose mothers
fact that F -isoprostanes are derived from free radical
were supplemented with fish oil or olive oil during pregnancy. Mean ±
oxidation of arachidonic acid, which is significantly
SEM. Between group differences were assessed using GLM. *P < 0.001
reduced following ω3 fatty acids. Therefore, the changes
after adjustment for red cell 20:4 ω6. Reproduced from Barden et al.42
in F -isoprostanes most likely reflect a true reduction in
oxidative stress, rather than as result of a reduction in thesupply of substrate.
independent of red cell 20:4 ω6 levels. Urinary F -iso-
How F -isoprostanes are reduced following ω3 fatty
prostanes corrected for creatinine excretion tended to be
acid supplementation remains unresolved. We suggested
lower in infants whose mother had taken fish oil (P = 0.06).
this might be explained, at least in part, by the anti-
Our findings are in accordance with several other studies
inflammatory effects of ω3 fatty acids and a reduction in
in which ω3 fatty acids have been supplemented. Quaggiotto
leukocyte free radical formation. Activated leukocytes
et al.43 showed that, compared to beef tallow, high doses of
generate powerful oxidants during phagocytosis46 and
ω3 fatty acids reduced plasma F -isoprostanes after coronary
cytokines such as TNF-α and IL-6 stimulate leukocytes
occlusion in pigs. Similarly, Higdon et al.25 demonstrated a
and endothelial cells to generate free radicals, further
fall in plasma F -isoprostanes in post-menopausal women
propagating the pro-oxidant condition. In support of this
given ω3 fatty acids compared with diets enriched in oleate
hypothesis, we showed that the changes in urinary F -
or linoleate. In the latter study, however, significant differ-
isoprostanes were significantly positively associated
ences were eliminated when F -isoprostanes were adjusted
with changes in TNF-α concentration.45 Numerous stud-
for plasma arachidonic acid concentrations.25
ies have demonstrated anti-inflammatory actions of ω3
We have demonstrated in two trials that both EPA and
fatty acids, with falls in cytokines most often observed
DHA equally reduced F -isoprostanes.44,45 In overweight,
following leukocyte stimulation.47 ω3-Fatty acids also
Published by Maney Publishing (c) W. S. Maney & Son Limited
Fig. 3. Change in urinary F -isoprostanes in (A) overweight, hyperlipidaemic men and (B) hypertensive Type 2 diabetic patients. Mean ± SEM. *P < 0.01
for a treatment effect (GLM). Reproduced from Mori et al.44,45 with permission.
have been shown to suppress production of reactive oxy-
Simopoulos AP, Kifer RR, Martin RE, Barlow SM. (eds) World
gen species (superoxide and hydrogen peroxide) in stim-
Review of Nutrition and Dietetics, vol. 66. Basel: Karger, 1991; 1–592.
Connor SL, Connor WE. Are fish oils beneficial in the prevention and
treatment of coronary artery disease? Am J Clin Nutr 1997; 66 (4
reduced in isolated human52 and rat54 polymorphonu-
Suppl): 1020S–1031S.
clear leukocytes, as well as in human monocytes.48,53
Mori TA, Burke V, Puddey IB et al. Purified eicosapentaenoic and
Other potential mechanisms may relate to the assembly
docosahexaenoic acids have differential effects on serum lipids and
of ω3 fatty acids in membrane lipids and lipoproteins
lipoproteins, LDL particle size, glucose, and insulin in mildlyhyperlipidemic men.
making the double bonds less susceptible to free radical
Mori TA, Bao DQ, Burke V, Puddey IB, Beilin LJ. Docosahexaenoic
attack,55 inhibition of the pro-oxidant enzyme phospholi-
acid but not eicosapentaenoic acid lowers ambulatory blood pressure
pase A ,56 and stimulation of antioxidant enzymes.57,58 In
this regard, there is evidence that ω3 fatty acids up-regu-
Mori TA, Watts GF, Burke V et al. Differential effects of
late gene expression of antioxidant enzymes and down-
eicosapentaenoic acid and docosahexaenoic acid on forearm vascularreactivity of the microcirculation in hyperlipidaemic, overweight men.
regulate genes associated with production of reactive
Nenseter MS, Drevon CA. Dietary polyunsaturates and peroxidation
Steinberg D, Parthasarathy S, Carew TE, Khoo JC, Witztum JL.
Beyond cholesterol. Modifications of low-density lipoprotein thatincrease its atherogenicity.
Liu J, Yeo HC, Doniger SJ, Ames BN. Assay of aldehydes from lipid
There is no evidence for a pro-oxidant effect of ω3 fatty
peroxidation: gas chromatography-mass spectrometry compared to
acids. Our findings and the recent literature demonstrate
that ω3 fatty acids do not adversely affect, and indeed
10. Visioli F, Colombo C, Galli C. Oxidation of individual fatty acids
may attenuate, oxidative stress. The results clearly high-
yields different profiles of oxidation markers.
light the need for caution in choosing methodologies for
11. Brude IR, Drevon CA, Hjermann I et al. Peroxidation of LDL from
the assessment of oxidative stress. Further studies are
combined-hyperlipidemic male smokers supplied with omega-3 fatty
also required to explore potential mechanisms for the
observation of an association between oxidative stress,
markers of inflammation and atherosclerosis following
12. Suzukawa M, Abbey M, Howe PR, Nestel PJ. Effects of fish oil fatty
acids on low density lipoprotein size, oxidizability, and uptake by
3 fatty acids. Nonetheless, there appears no reason why
3 fatty acids should not be taken either as fish meals or
13. Wander RC, Du SH, Ketchum SO, Rowe KE. Effects of interaction of
fish oils capsules, in view of their overall favourable
RRR-alpha-tocopheryl acetate and fish oil on low-density-lipoprotein
effects on cardiovascular risk reduction.
oxidation in postmenopausal women with and without hormone-replacement therapy
14. Hau MF, Smelt AH, Bindels AJ et al. Effects of fish oil on oxidation
resistance of VLDL in hypertriglyceridemic patients. ArteriosclThromb Vasc Biol 1996; 16): 1197–1202.
15. Lussier-Cacan S, Dubreuil-Quidoz S, Roederer G et al. Influence of
The studies described were supported by grants from the
probucol on enhanced LDL oxidation after fish oil treatment of
National Health and Medical Research Council of
Australia, the West Australian Health Promotion
16. Harats D, Dabach Y, Hollander G et al. Fish oil ingestion in smokers
Foundation (Healthway) and the Royal Perth Hospital
Published by Maney Publishing (c) W. S. Maney & Son Limited
and nonsmokers enhances peroxidation of plasma lipoproteins
Medical Research Foundation. Purified eicosapen-
taenoic and docosahexaenoic acids and olive oil cap-
17. Stalenhoef AF, de Graaf J, Wittekoek ME, Bredie SJ, Demacker PN,
sules were kindly provided by the Fish Oil Test
Kastelein JJ. The effect of concentrated n-3 fatty acids versusgemfibrozil on plasma lipoproteins, low density lipoprotein
Materials Program and the US National Institutes of
heterogeneity and oxidizability in patients with hypertriglyceridemia.
Health. I would like to acknowledge my collaborators,
Assoc. Prof. Kevin Croft, Prof. Ian Puddey, Prof. Lawrie
18. Higgins S, Carroll YL, McCarthy SN et al. Susceptibility of LDL to
Beilin, Dr Valerie Burke, Dr Anne Barden, Assoc. Prof.
oxidative modification in healthy volunteers supplemented with low
Susan Prescott, Dr David Dunstan, Dr Richard
doses of n-3 polyunsaturated fatty acids.
19. Bonanome A, Biasia F, De Luca M et al. n-3 fatty acids do not
Woodman and Dr Jan Dunstan, and the technical assis-
enhance LDL susceptibility to oxidation in hypertriacylglycerolemic
20. Nenseter MS, Rustan AC, Lund-Katz S et al. Effect of dietary
supplementation with n-3 polyunsaturated fatty acids on physicalproperties and metabolism of low density lipoprotein in humans.
21. Frankel EN, Parks EJ, Xu R, Schneeman BO, Davis PA, German JB.
Mori TA, Beilin LJ. ω3 Fatty acids, blood lipids and cardiovascular
Effect of n-3 fatty acid-rich fish oil supplementation on the oxidation
Effect of fish and fish oil-derived omega-3 fatty acids on lipid oxidation
22. Bittolo-Bon G, Cazzolato G, Alessandrini P, Soldan S, Casalino G,
40. Pratico D. F -isoprostanes: sensitive and specific non-invasive indices
Avogaro P. Effects of concentrated DHA and EPA supplementation on
LDL peroxidation and vitamin E status in type HB hyperlipidemic
41. Mori TA, Dunstan DW, Burke V et al. Effects of dietary fish and
patients. In: Drevon CA, Bakaas I, Krokan HE. (eds) Omega-3 Fatty
exercise training on urinary F -isoprostane excretion in non-insulin
Acids. Basel: Birkhauser, 1993; 51–58.
23. Devaraj S, Hirany SV, Burk RF, Jialal I. Divergence between LDL
42. Barden AE, Mori TA, Dunstan JA et al. Fish oil supplementation in
oxidative susceptibility and urinary F2-isoprostanes as measures of
pregnancy lowers F -isoprostanes in neonates at high risk of atopy. Free Radic Res 2004, 38: 233–239.
24. Halliwell B. Oxidative stress, nutrition and health. Experimental
43. Quaggiotto P, Leitch JW, Falconer J, Murdoch RN, Garg ML. Plasma
strategies for optimization of nutritional antioxidant intake in humans.
F isoprostane levels are lowered in pigs fed an (n-3) polyunsaturated
fatty acid supplemented diet following occlusion of the left anterior
25. Higdon JV, Liu JK, Du SH, Morrow JD, Ames BN, Wander RC.
Supplementation of postmenopausal women with fish oil rich in
44. Mori TA, Puddey IB, Burke V et al. Effect of ω3 fatty acids on
eicosapentaenoic acid and docosahexaenoic acid is not associated with
oxidative stress in humans: GCMS measurement of urinary F2-
greater in vivo lipid peroxidation compared with oils rich in oleate and
linoleate as assessed by plasma malondialdehyde and F -isoprostanes.
45. Mori TA, Woodman RJ, Burke V, Puddey IB, Croft KD, Beilin LJ.
Effect of eicosapentaenoic acid and docosahexaenoic acid on
26. Wander RC, Du SL. Oxidation of plasma proteins is not increased
oxidative stress and inflammatory markers, in treated-hypertensive
after supplementation with eicosapentaenoic and docosahexaenoic
46. Babior BM. Phagocytes and oxidative stress.
27. Allard JP, Kurian R, Aghdassi E, Muggli R, Royall D. Lipid
peroxidation during n-3 fatty acid and vitamin E supplementation in
47. Calder PC. N-3 polyunsaturated fatty acids, inflammation and
28. Palozza P, Sgarlata E, Luberto C et al. n-3 fatty acids induce oxidative
modifications in human erythrocytes depending on dose and duration
48. Hiramatsu K, Arimori S. Increased superoxide production by
mononuclear cells of patients with hypertriglyceridemia and diabetes.
29. Hansen JB, Berge RK, Nordoy A, Bonaa KH. Lipid peroxidation of
isolated chylomicrons and oxidative status in plasma after intake of
49. Thompson PJ, Misso NL, Passarelli M, Phillips MJ. The effect of
eicosapentaenoic acid consumption on human neutrophil
30. Ando K, Nagata K, Beppu M et al. Effect of n-3 fatty acid
50. Varming K, Schmidt EB, Svaneborg N et al. The effect of n-3 fatty
supplementation on lipid peroxidation and protein aggregation in rat
31. Uusitupa MI, Niskanen L, Luoma J et al. Autoantibodies against
51. Fisher M, Levine PH, Weiner BH et al. Dietary n-3 fatty acid
oxidized LDL do not predict atherosclerotic vascular disease in non-
supplementation reduces superoxide production and
chemiluminescence in a monocyte-enriched preparation of leukocytes.
32. Hafeman DG, Hoekstra WG. Lipid peroxidation in vivo during
52. Fisher M, Upchurch KS, Levine PH et al. Effects of dietary fish oil
vitamin E and selenium deficiency in the rat as monitored by ethane
supplementation on polymorphonuclear leukocyte inflammatory
evolution. J Nutr 1977; 107: 666–672.
33. Mabile L, Piolot A, Boulet L et al. Moderate intake of n-3 fatty acids
53. Sirtori CR, Gatti E, Tremoli E et al. Olive oil, corn oil, and n-3 fatty
is associated with stable erythrocyte resistance to oxidative stress in
acids differently affect lipids, lipoproteins, platelets, and superoxide
formation in type II hypercholesterolemia.
34. Sanders TA, Hinds A. The influence of a fish oil high in
docosahexaenoic acid on plasma lipoprotein and vitamin E
54. Carbonell T, Rodenas J, Miret S, Mitjavila MT. Fish oil and oxidative
concentrations and haemostatic function in healthy male volunteers.
Published by Maney Publishing (c) W. S. Maney & Son Limited
35. Haglund O, Luostarinen R, Wallin R, Wibell L, Saldeen T. The effects
55. Applegate KR, Glomset JA. Computer-based modeling of the
of fish oil on triglycerides, cholesterol, fibrinogen and
conformation and packing properties of docosahexaenoic acid.
malondialdehyde in humans supplemented with vitamin E. J Nutr
1991; 121: 165–169.
56. von Schacky C, Siess W, Fischer S, Weber PC. A comparative study
36. Brown JE, Wahle KW. Effect of fish-oil and vitamin E
of eicosapentaenoic acid metabolism by human platelets in vivo and in
supplementation on lipid peroxidation and whole-blood aggregation in
57. Demoz A, Willumsen N, Berge RK. Eicosapentaenoic acid at
37. Meydani M, Natiello F, Goldin B et al. Effect of long-term fish oil
hypotriglyceridemic dose enhances the hepatic antioxidant defense in
supplementation on vitamin E status and lipid peroxidation in women. J Nutr 1991; 121: 484–491.
58. Olivieri O, Negri M, De Gironcoli M et al. Effects of dietary fish oil
38. Fam SS; Morrow JD. The isoprostanes: unique products of
on malondialdehyde production and glutathione peroxidase activity in
59. Takahashi M, Tsuboyama-Kasaoka N, Nakatani T et al. Fish oil
39. Morrow JD. The isoprostanes: their quantification as an index of
feeding alters liver gene expressions to defend against PPAR alpha
activation and ROS production. Am J Physiol 2002; 282: G338–G348.
International Journal of Pharmacognosy and Phytochemical Research 2013; 5(2); 113-119Gastroprotective Efficacy of Folic Acid and Omeprazole in1Samar Morjan , *1Shaza Al Laham , 2Rana Atieh1Pharmacology & Toxicology Department - Faculty of pharmacy- Damascus University- Damascus-Syria2Histopathological Department - Medicin faculty- Damascus University- Damascus-Syria ABSTRACT Gastric and
Dechra Veterinary Products Limited (A business unit of Dechra Pharmaceuticals PLC) Sansaw Business Park Hadnall, Shrewsbury Shropshire SY4 4AS Tel: 01939 211200 BOVOCYCLINE PESSARY 2000 MG damage to the liver. Photodermatitis often occurs in INTRAUTERINE PESSARY FOR CATTLE areas of sparsely pigmented skin if these are exposedto sunlight. Allergic reactions are rare. In case of Qualitat
 gas chromatography mass spectrometry. In addition,plasma TBARS were significantly elevated following
ω3 fatty acids, whereas MDA was reduced. Whenplasma MDA levels were normalized to plasma polyun-saturated fatty acid concentrations, significant differ-ences were eliminated.25
Other measures of oxidative stress include electron
spin resonance detecting free radical species,24 measure-ment of antibodies to oxidized LDL31 and breath excre-tion of ethane and pentane.24 The latter assay, however,has yielded variable results in animals32 and humans27supplemented with ω3 fatty acids. Other literaturereports have shown that ω3 fatty acids had no adverseeffects on plasma protein oxidation26 and rendered ery-throcytes more resistant to haemolysis following oxida-tive challenge.33
On the basis of studies reporting adverse effects on
Fig. 1. Change in urinary F -isoprostane excretion from baseline to post-
gas chromatography mass spectrometry. In addition,plasma TBARS were significantly elevated following
ω3 fatty acids, whereas MDA was reduced. Whenplasma MDA levels were normalized to plasma polyun-saturated fatty acid concentrations, significant differ-ences were eliminated.25
Other measures of oxidative stress include electron
spin resonance detecting free radical species,24 measure-ment of antibodies to oxidized LDL31 and breath excre-tion of ethane and pentane.24 The latter assay, however,has yielded variable results in animals32 and humans27supplemented with ω3 fatty acids. Other literaturereports have shown that ω3 fatty acids had no adverseeffects on plasma protein oxidation26 and rendered ery-throcytes more resistant to haemolysis following oxida-tive challenge.33
On the basis of studies reporting adverse effects on
Fig. 1. Change in urinary F -isoprostane excretion from baseline to post-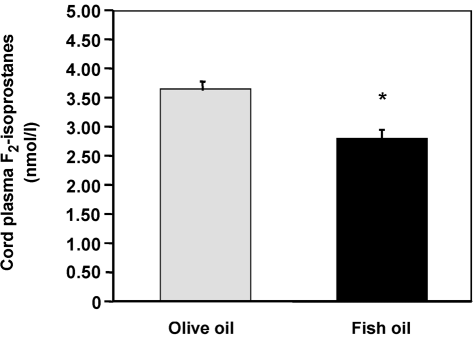
 Effect of fish and fish oil-derived omega-3 fatty acids on lipid oxidation
mildly hyperlipidaemic men, supplementation with 4 gdaily of purified EPA or DHA for 6 weeks decreased post-intervention urinary F -isoprostane levels by 27% follow-
ing EPA (1.24 nmol/24-h, P < 0.0001) and 26% followingDHA (1.20 pmol/24-h, P < 0.0001), relative to an olive oilcontrol group, after adjusting for baseline values (Fig.
Effect of fish and fish oil-derived omega-3 fatty acids on lipid oxidation
mildly hyperlipidaemic men, supplementation with 4 gdaily of purified EPA or DHA for 6 weeks decreased post-intervention urinary F -isoprostane levels by 27% follow-
ing EPA (1.24 nmol/24-h, P < 0.0001) and 26% followingDHA (1.20 pmol/24-h, P < 0.0001), relative to an olive oilcontrol group, after adjusting for baseline values (Fig.