Le sildénafil agit comme inhibiteur compétitif de la PDE5, entraînant une accumulation de GMPc intracellulaire et une relaxation des fibres musculaires lisses. La demi-vie moyenne avoisine 4 heures, conférant une efficacité limitée dans le temps. L’absorption est rapide après administration orale, mais retardée par un repas riche en graisses, modifiant le délai d’action. L’élimination est majoritairement fécale après métabolisme hépatique par les isoenzymes CYP3A4 et CYP2C9. Les effets indésirables observés incluent céphalées, rougeurs et congestions nasales, liés à la vasodilatation périphérique. Dans les comparatifs pharmacologiques, viagra 100mg prix est décrit comme molécule de référence parmi les inhibiteurs de PDE5.
Rheinischesmuseumfuerphilologie.de
LE QUÉRÉ, A., D. P. WRIGHT, B. SÖDERSTRÖM, A. TUNLID
corrhizal fungi and adventitious root formation in Scots
and T. JOHANSSON (2005): Global patterns of gene regu-
pine in vitro. Tree Physiol 22: 373–381.
lation associated with the development of ectomycor-
NIEMI, K., M. SALONEN, A. ERNSTSEN, H. HEINONEN-TANSKI
rhiza between birch (Betula pendula Roth.) and Paxil-
and H. HÄGGMAN (2000): Application of ectomycorrhizal
lus involutus (Batsch) Fr. Mol Plant-Microbe Interact
fungi in rooting of Scots pine fascicular shoots. Can J
18: 659–673.
For Res 30: 1221–1230.
LI, L., S. LU and V. CHIANG (2006): A genomic and molecu-
NIEMI, K., C. SCAGEL and H. HÄGGMAN (2004): Application
lar view of wood formation. Crit Rev Plant Sci 25:
of ectomycorrhizal fungi in vegetative propagation of
conifers. Plant Cell Tissue Organ Cult 78: 83–91.
LINDAHL, B. O., A. F. S. TAYLOR and R. D. FINLAY (2002):
PILATE, G., E. GUINEY, K. HOLT, M. PETIT-CONIL,
Defining nutritional constraints on carbon cycling in
C. LAPIERRE, J.-C. LEPLÉ, B. POLLET, I. MILA, E. A. WEB-
boreal forests – towards a less ‘phytocentric’ perspec-
STER, H. G. MARSTORP, D. W. HOPKINS, L. JOUANIN,
tive. Plant Soil 242: 123–135.
W. BOERJAN, W. SCHUCH, D. CORNU and C. HALPIN
(2002): Field and pulping performances of transgenic
LOYD, G. and B. MCCOWN (1980): Commercially-feasible
micropropagation of Mountain Laurel, Kalmia latifolia,
trees with altered lignification. Nat Biotechnol 20:
by use of shoot-tip culture. Int Plant Prop Soc Proc 30:
ROLANDO, C., B. MONTIES and C. LAPIERRE (1992): Thioaci-
dolysis, pp 334–349. In: Methods in lignin chemistry,
MARX, D. H. (1969): The influence of ectotrophic fungi on
edited by LIN, S. Y. and C. W. DENCE, Springer-Verlag,
the resistance of pine roots to pathogenic infections. I.
Antagonism of mycorrhizal fungi to root pathogenic
SARKANEN, K. V. and H. L. HERGERT (1971): Classification
fungi and soil bacteria. Phytopathology 59: 153–163.
and distribution, pp 43–94. In: Lignins: Occurrence,
MOREL, M., C. JACOB, A. KOHLER, T. JOHANSSON,
formation, structure and reaction, edited by SARKANEN,
F. MARTIN, M. CHALOT and A. BRUN (2005): Identifica-
K. V. and C. H. LUDWIG, Wiley-Interscience, New York.
tion of genes differentially expressed in extraradical
SEPPÄNEN, S.-K., H.-L. PASONEN, S. VAURAMO, J. VAHALA,
mycelium and ectomycorrhizal roots during Paxillus
M. TOIKKA, I. KILPELÄINEN, H. SETÄLÄ, T. H. TEERI,
involutus-Betula pendula ectomycorrhizal symbiosis.
S. TIMONEN and A. PAPPINEN (2007): Decomposition of
Appl Environ Microbiol 71: 382–391.
the leaf litter and mycorrhiza forming ability of silver
MORIN, C., J. SAMSON and M. DESSUREAULT (1999): Protec-
birch with a genetically modified lignin biosynthesis
tion of black spruce seedlings against Cylindrocladium
pathway. Appl Soil Ecol 36: 100–106.
root rot with ectomycorrhizal fungi. Can J Bot 77:
SMITH, S.E. and D. J. READ (1997): Mycorrhizal symbiosis.
NIEMI, K. and H. HÄGGMAN (2002): Pisolithus tinctorius
TIIMONEN, H., T. ARONEN, T. LAAKSO, P. SARANPÄÄ, V. CHI-
promotes germination and forms mycorrhizal structures
ANG, T. YLIOJA, H. ROININEN and H. HÄGGMAN (2005):
in Scots pine somatic embryos in vitro. Mycorrhiza 12:
Does lignin modification affect feeding preference or
growth performance of insect herbivores in transgenic
NIEMI, K., H. HÄGGMAN and T. SARJALA (2002): Effects of
Roth)? Planta 222:
exogenous diamines on the interaction between ectomy-
Genetic Diversity of the Relict Plant Taiwania cryptomerioides Hayata (Cupressaceae) in Mainland China
By ZHONG-CHAO LI1), XIAO-LAN WANG2) and XUE-JUN GE1),*)
Abstract
repeats (ISSR). In comparison with other coniferous
The genetic diversity and differentiation of five popu-
species, T. cryptomerioides from mainland China pos-
lations of Taiwania cryptomerioides Hayata in mainland
sesses little genetic variation, particularly at the level of
China were investigated using inter-simple sequence
individual populations (the percentage of polymorphicloci, Nei’s gene diversity and Shannon’s indices of diver-
sity at the species and population levels are 38.02 %,
) South China Botanical Garden, The Chinese Academy of
0.1326, 0.1986 and 9.27 %, 0.035, 0.0518 respectively).
Sciences, Guangzhou 510650, P. R. China.
In contrast, the level of population differentiation is
) Center for Functional Genomics and Microarray, Guangzhou
much higher (G : 0.7269; Shannon’s genetic differentia-
θ B: 0.668; AMOVA genetic differen-
) Corresponding author: Dr. XUE-JUN GE. South China Botanical
tiation: 72.37 %). The genetic divergence of pairs of pop-
Garden, The Chinese Academy of Sciences, Guangzhou 510650,P. R. China. Tel: +86-20-3725 2551; Fax: +86-20-3725-2831. E-
ulations was not significantly correlated with the geo-
mail address: [email protected], [email protected]
graphical distance separating them. Current patterns of
genetic variation were related to biogeographic history
Information about the genetic structure of a rare
and the small population size. On the basis of these
species such as this, as well as data on its overall level
findings, we discuss the development of conservation
of genetic diversity, are particularly important for
strategies for this endangered species.
species conservation. Genetic analyses can provide valu-able insights into the processes influencing extinction
Key words: Taiwania cryptomerioides; genetic diversity, ISSR,China, relict species, conservation
(CLARKE and YOUNG, 2000) and, increasingly, geneticdata have been used to define conservation managementunits and to predict changes in population structure and
Introduction
dynamics (NEWTON et al., 1999). However, in comparison
Like most of the genera within the Cupressaceae that
with the rarities from Europe and North America, few
evolved since the Jurassic period, Taiwania, one of the
detailed studies of genetic variation have been under-
so-called “living fossils”, is considered to be an ancient
taken on rare plants in China. Subtropical mainland
genus in evolutionary terms with its origin dating back
Asia was one of the most important refugia for plants
to the Tertiary. According to the fossil evidence, e.g. Tai-
during the Pleistocene glaciations because it was not
wania schaeferi Schloem.-Jäg. discovered in Klein-
covered by ice sheets. Many species that became extinct
saubernitz (Germany) (WALTHER, 1999), Spitsbergen
elsewhere survived in this region, but in a state of isola-
(Norway) and Alaska (LePage BA, unpublished data),
tion and in small populations. Strong genetic differenti-
this genus was widespread in what are now the temper-
ation has been discovered in some relict conifer species
ate regions of the northern hemisphere (FLORIN, 1963).
in China, for example Cathaya argyrophylla (GE et al.,
Today, Taiwania, a monotypic genus containing only
1998), Glyptostrobus pensilis (LI and XIA, 2005), Metase-Taiwania cryptomerioides Hayata, has a scattered dis-
quoia glyptostroboides (LI et al., 2005) and Ametotaxus
tribution. Its distributional range covers mountain
argotaenia (GE et al., 2005). The genetic diversity of
areas on the border between Burma and China (south-
T. cryptomerioides from Taiwan has been assessed previ-
western part of Yunnan province), the Hoang Lien Son
ously by allozyme (LIN et al., 1993), ISSR and ITS
mountain range in northern Vietnam, and Taiwan
sequence analysis (CHANG, 2005). A low genetic differen-
(FARJON, 2002; FARJON and GARCIA, 2003). Taiwan is an
tiation (G : 5.2 % based on allozyme data, LIN et al.,
important refuge for T. cryptomerioides: it grows on the
1993; Φ : 15.26 % using ISSR, CHANG, 2005) was found
Central Mountain at the altitudes of 1600–2600 m,
in the Taiwanese populations. Nevertheless, the
where the largest population and greatest timber yield
intraspecific genetic variation across the distributional
in the world are found (HUANG, 1983). In addition, there
are still a few individuals of T. cryptomerioides in main-
Among various molecular tools, the inter-simple
land China: in the remote Lichuan mountains in Hubei
sequence repeats (ISSR) method has been widely used
Province, Leishan mountains in Guizhou Province, and
for studies of population genetics, because the repeats
Gutian-Pingnan mountains in Fujian Province. These
are highly variable, and the method is economical in
areas are regarded as having acted as key refugia for
terms of time, money, and labor (GUPTA et al., 1994;
relict species during Pleistocene glaciations (WU, 1980;
ZIETKIEWICZ et al., 1994; TSUMURA et al., 1996; WOLFE
WANG and LIU, 1994). The populations in mainland
and LISTON, 1998). ISSRs have also been used to deter-
China have only been widely known to botanists since
mine the genetic diversity of species of conservation con-
the early decades of the 20th century, and have some-
cern (ESSELMAN et al., 1999). Technically, the ISSR reac-
times been regarded as comprising a different species,
tion is more specific than RAPD amplification due to the
namely T. flousiana Gaussen. Due to human overex-
longer SSR-based primers (WOLFE and LISTON, 1998).
ploitation in the past century, the conservation status of
One limitation of the ISSR technique is that the bands
this species is “vulnerable” according to the IUCN Red
are scored as dominant markers and that genetic diver-
List criteria (FARJON, 2001) and it is a protected species
sity estimates are based on diallelic characters, thus the
level of genetic diversity may be underestimated. The
T. cryptomerioides is a component of evergreen
primary objective of this study was to use ISSR markers
broadleaf forests in mountainous areas. It occurs at an
to investigate genetic variation within and between the
altitude of 500–2800 m in “coniferous, broad-leaved, or
extant T. cryptomerioides populations in mainland
mixed evergreen valley forests on acid, red, or brown
China. We wished to determine whether these popula-
soils in warm or warm temperate regions with high
tions are strongly differentiated, as is the case for the
summer and autumn rainfall but drier winters, usually
other relict conifer species mentioned above. The result
scattered and associated with Chamaecyparis formosen-
of this investigation could assist in the development of
sis, C. obtusa var. formosana, Cunninghamia lanceolata,
conservation strategies, by identifying those populations
Pinus wallichiana, or Tsuga dumosa, but sometimes
of particular importance in terms of their genetic char-
forming pure stands” (FU et al., 1999). It can grow as
tall as 70 meters and is therefore known as the ‘King ofAsian Conifers’. This diploid species (2n = 22, LI et al.,
Material and Methods
2000) is wind-pollinated and monoecious, with male andfemale cones occurring on different branches (FU et al.,
1999). With two subapical, unequal wings, seeds of
A total of 104 individuals of Taiwania cryptomerioidesT. cryptomerioides are light and small, and can be car-
was studied, representing four extant wild populations
in mainland China and one plantation population (TC)
Table 1. – Genetic variability within populations of Taiwania cryptomerioides revealed by ISSR (N: sample size per population; P: percentage of polymorphic loci; H : Nei’s gene diversity; Hpop: Shannon indices).
(Table 1, Fig. 1). Among the four wild populations, many
were carried out in a PTC-200 thermal cycler (MJ
individuals are several hundred years old with a DBH
research) following the method of GE and SUN (1999).
greater than 80 cm; their ecological and phytocoenologi-
One hundred primers, each 15–23 nucleotides long
cal features have been described previously (Table 1;
(UBC primer set # 9, Biotechnology Laboratory, Univer-
QIU et al., 1984; LIU et al., 1987; GE, 1995; FENG et al.,
sity of British Columbia), were screened for the produc-
1998; YANG and JIANG, 1999). Individuals were sampled
tion of a high proportion of polymorphic and repro-
at random, regardless of their size and age. Young,
ducible banding. Fifteen primers (UBC # 807, 808, 811,
healthy leaves were collected in the field, and dried in
826, 827, 828, 835, 836, 840, 846, 849, 855, 866, 880, and
silica gel prior to DNA extraction. Total DNA was
886) fulfilled these requirements. PCR products were
extracted using the CTAB method described by DOYLE
electrophoresed on 2.0 % agarose gels buffered with 0.5 x
(1991) then dissolved in 100 µL of TE buffer. DNA con-
TBE. A 100 bp DNA Ladder (New England Biolabs) was
centration was determined by comparison to uncut
used as a size marker. After staining with ethidium bro-
lambda DNA on 1% agarose gels; the average DNA con-
mide, the fragments were identified using image analy-
centration was about 20–30 ng/µl.
sis software for gel documentation (LabWorks SoftwareVersion 3.0; UVP, Upland, CA 91786, USA).
PCR amplification was carried out in a volume of
20 µl; the reaction comprised 20 ng of template DNA,
Only bands that could be unambiguously interpreted
10mM Tris-HCl (pH 8.0), 50mM KCl, 0.1% Triton x 100,
across all the population samples were used in this
2.5mM MgCl , 0.1mM dNTPs, 2 % formamide, 0.2 µM
study. ISSR profiles were scored as discrete characters
primer and 1.5 units of Taq polymerase. Amplifications
(presence or absence of amplified products) for each
Figure 1. – The sampling locations of Taiwania cryptomerioides Hayata.
individual. The resulting data matrix was analyzed
were 0.0518 at the population level (Hpop) and 0.1986
using POPGENE v. 1.31 (YEH et al., 1999) to estimate
at the species level (Hsp), respectively (Table 1). The cul-
genetic diversity parameters: the percentage of polymor-
tivated TC population exhibited the lowest genetic
phic loci (P) and expected heterozygosity (H ). At the
diversity. Among the four wild populations, population
species level, genetic diversity measures (H : total gene
GP had the greatest level of variability (P: 20.31%, H :
diversity; G : coefficient of gene differentiation) and the
0.0806, Hpop: 0.1179); population LS had the lowest
level of gene flow (Nm) were measured using Nei’s
level of variability (P: 3.65%, H : 0.0121, Hpop: 0.0186)
(1973) gene diversity statistics. Genetic diversity was
(Table 1). The average polymorphism of the four wild
also estimated using Shannon’s information statistic
= -Σpi log p where p is the fre-
The coefficient of genetic differentiation between pop-
ulations (G ) was 0.7269, estimated by partitioning the
two levels: the average diversity within the populations
total gene diversity. The Shannon’s diversity index
(Hpop), and the total diversity (Hsp). Then the propor-
analysis partitioned 73.92 % of the total variation
tion of diversity between the populations was estimated
between populations, in broad agreement with the
as: (Hsp-Hpop)/Hsp.
result of genetic differentiation analysis. The level of
In addition, an analysis of molecular variance
gene flow (Nm) was estimated to be 0.0939. The AMOVA
(AMOVA; EXCOFFIER et al., 1992) was used to estimate
analysis provided corroborating evidence for the genetic
variance components for ISSR phenotypes, partitioning
structure obtained from Nei’s genetic diversity statistics
the variation between populations and between individ-
and Shannon’s diversity estimate. There were highly
uals. AMOVA analysis was performed using the Arle-
significant (P < 0.001) genetic differences between the
quin 2.000 program (SCHNEIDER et al., 2000). To test the
five populations of T. cryptomerioides. Of the total mole-
presence of isolation by distance, a Mantel test between
cular variance, 72.37 % was attributable to between-pop-
the pairwise genetic distance and geographic distance
ulation diversity and the rest (27.63 %) to differences
from each population pair was applied using Mantel’s
between individuals within populations (Table 2). A sim-
test in the software TFPGA (MILLER, 1997) (1000 per-
ilar result was obtained from the Hickory calculation: θ B
mutations). Pairwise genetic distances (Φ ) were got
was 0.668 for the f free model, which had the smallest
from AMOVA analysis, which are analogous to tradition-
Genetic distances (Φ ) between populations of
An alternative Bayesian method, allowing direct esti-
T. cryptomerioides were calculated by Arlequin 2.000
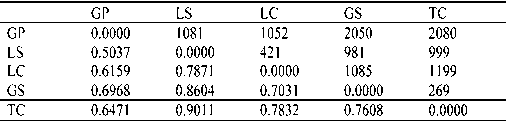
(SCHNEIDER et al., 2000) (Table 3). The highest distance
ous knowledge of F , was employed to estimate θ B
value was 0.9011 between TC and LS, and the lowest
(analogous to F ) using Hickory 1.0 (HOLSINGER and
was 0.5037 between LS and GP. Geographically TC and
LEWIS, 2003). Using the default sampling parameters
GS are the closest pair of sites (269 km), however, the
(burn-in = 5 000, sample = 25 000, thin = 5), θ B was cal-
genetic distance between them, 0.7608, was not the low-
culated under four different models. The first was a full
est of all the pairs. The result of a Mantel test with 1000
model, in which both θ B and f (i.e., F , the inbreeding
permutations revealed that the genetic divergence of
coefficient) were estimated. Two alternative models
populations was not significantly correlated with geo-
assumed either θ B or f equal to zero. Finally, because
graphical distance (Mantel test, r = -0.3124, P = 0.2170).
estimates of f based on dominant markers are usuallystrongly biased (especially at low sample sizes, i.e.,n < 10), HICKORY was used to construct a model in
Discussion
which f was allowed to vary. The deviance information
Low level of genetic diversity and strong genetic differen-
criterion (DIC) was employed to determine which model
was the best fit for the data: lower values indicate a bet-
Correlations between genetic diversity and various
attributes of different species have been examined based
Table 3. – The pairwise genetic distance (ΦST) (below diagonal)
In this study, 73 of the 192 electrophoretic bands
and geographic distance (km) (above diagonal).
(38.02 %) were polymorphic within the populations. Thepercentages of polymorphic loci (P) for a single popula-tion ranged from 1.56 % (TC) to 20.31% (GP), with anaverage of 9.27 %. The average genetic diversity wasestimated to be 0.0350 at the population level (H ) and
0.1282 at the species level (H ). The Shannon indices
Table 2. – Analysis of molecular variance (AMOVA) for five populations of Taiwania cryptomerioidesd.f.: degrees of freedom; SS: sum of squares; MS: expected mean squares.
a Significance tests after 1000 permutations.
on a vast amount of data relating to natural plant popu-
pensilis (LI and XIA, 2005), Metasequoia glyptostroboides
lations (HAMRICK and GODT, 1989; NYBOM, 2004). Most
(LI et al., 2005), Cupressus chengiana (HAO et al., 2006),
conifers have high levels of genetic diversity and low
Torreya jackki (LI and JIN, 2007) and Abies ziyuanensis
levels of differentiation between populations (HAMRICK
(TANG et al., 2007) (Table 4). Since these species are
et al., 1992). Examples include Fitzroya cupressoides
characterized by their relict status, biogeographic histo-
(Hpop: 0.547, Φ : 0.1438; ALLNUTT et al., 1999), Junipe-
ry may play an important role in determining their
rus rigida, J. coreana (H : 0.224 and 0.199; G : 0.173
genetic diversity. T. cryptomerioides is one of the repre-
and 0.118, respectively; HUH and HUH, 2000), Cedrus
sentatives of relict gymnosperms from the Tertiary peri-
atlantica (Nei’s gene diversity within population, H :
od that are found in China. Fossil evidence indicates
0.194; F : 0.148; θ B: 0.178; RENAU-MORATA et al., 2005)
that the ancestor of T. cryptomerioides was much more
and Juniperus phoenicea (H : 0.148, H : 0.130, G :
widespread during the Tertiary than its current distrib-
0.12; MELONI et al., 2006). Similar genetic diversity and
ution suggests (WALTHER, 1999). Accordingly, a reason-
genetic differentiation have also been reported for the
able hypothesis is that the modern range of T. cryptome-
Taiwanese populations of T. cryptomerioides (LIN et al.,
rioides was the result of population fragmentation and
1993; CHANG, 2005). In this study, however, ISSR poly-
contraction after the Quaternary glacial cycles. The
morphisms revealed a low level of genetic diversity in
remnant populations contained only part of the genetic
the mainland populations of T. cryptomerioides, with an
variation present before the range reduction. Genetic
average of only 11.19% of ISSR bands being polymorphic
drift and inbreeding may have further decreased the
in the four wild populations studied. In addition, strong
genetic diversity and increased genetic differentiation in
population differentiation was demonstrated based on
the small, isolated populations of T. cryptomerioides in
Nei’s gene diversity, Shannon’s information statistic,
AMOVA and Bayesian estimates. About 70 % of the total
In addition to historical influences, the present small
genetic variation was partitioned between the popula-
population size of T. cryptomerioides may be important.
tions of T. cryptomerioides studied in mainland China.
According to the literature there are currently about
The low genetic diversity within populations and high
6000–7000 T. cryptomerioides individuals in mainland
genetic differentiation among populations we observed
China (HU et al., 1995); populations LC and GP are com-
in T. cryptomerioides are similar to that of many relict-
posed of only a dozen individuals (GE, 1995; YANG et al.,
ual endangered conifer species in mainland China.
2006). This suggests that this species is experiencing
These include Cathaya argyrophylla (GE et al., 1998),
erosion of its genetic diversity. In this study, only four-
Ametotaxus argotaenia (GE et al., 2005), Glyptostrobus
teen electrophoretic bands (8 %) with a frequency lower
Table 4. – Our results compared with those of relict gymnosperm species endemic to China. P: percentage of polymorphic loci; H : Nei’s gene diversity; Hpop: Shannon indices; G : the coefficient of gene differentiation.
than 30 % were observed, suggesting a possible stochas-
ies, which can be used to determine a species’ natural
tic process of genetic drift, which could result in the loss
range (BEEBEE and ROWE, 2004). Further study is need-
of low-frequency alleles in the populations (BROYLES,
ed to screen out the mtDNA fragments with intraspecific
polymorphic variation in this species.
The genetic structure of plant populations reflects the
interactions of various evolutionary processes. Low lev-
Conservation Implications
els of genetic differentiation in gymnosperms are usual-ly attributed to wind-pollination and breeding systems
The maintenance of genetic diversity is crucial to the
that promote outcrossing. T. cryptomerioides is dioe-
survival of organisms, because it allows them to evolve
cious, wind pollinated and has light seeds that are usu-
and adapt to changing environmental conditions. The
ally dispersed by wind (about 1000 seeds in 1.01 g;
high level of genetic differentiation between the studied
populations of Taiwania cryptomerioides indicates that
HUNG and CHANG, 1990). It seems likely, therefore, that
considerable gene flow should take place between popu-
a considerable amount of the overall genetic variation of
lations. For Taiwan populations of T. cryptomerioides,
the species in mainland China would be lost if manage-
little genetic differentiation is apparent, thus support-
ment concentrated only on the remaining large popula-
tions. Therefore, several populations throughout the
geographic proximity of the populations in Taiwan, the
entire range should be considered for conservation. At
populations in mainland China are separated from each
present three populations in mainland China (GS, LS
other by several hundred kilometers (Table 3). This
and LC) are protected in situ. Despite being situated
extreme isolation makes gene flow negligible, resulting
within a protected area, population LC has been severe-
in the lack of a correlation between genetic distance and
ly affected by land reclamation, and should therefore be
geographical distance found in this study. For example,
accorded a high priority for future conservation action.
the genetic distance between populations that are rela-
As for ex situ conservation in botanical gardens, further
tively close geographically (i.e., populations TC and GS)
reduction of genetic variability by genetic drift should be
was no lower than between geographically more distant
avoided, therefore one additional measure could be arti-
ones. The present locations of T. cryptomerioides in
ficial gene flow between populations, i.e., artificial intro-
mainland China coincide with the key refugia for other
ductions should include as many populations as possi-
relict species during the Pleistocene glaciations (W
1980; WANG and LIU, 1994). For example, at the north-ern limit of this species, Lichuan (LC) in Hubei
Acknowledgments
Province, there are many relict species, including
We thank Mr. GUO-SHENG HE, YONG-FU YU and YONG-
Davidia involucrata, Ginkgo biloba, Keteleeria fortunei,
MING YUAN for their help in collecting samples. This work
Taxus chinensis, Tetracentron sinensis, Emmenopterys
was supported by the National Natural Science Founda-
henryi and Bretschneidera sinensis. The famous relic
plant Metasequoia glyptostroboides was also discoveredin this region in the 1940s (see http://www.metase-
References
quoia.org/). This distribution pattern indicates that pop-ulations of T. cryptomerioides in mainland China may
ALLNUTT, T. R., A. C. NEWTON, A. LARA, A. PREMOLI,
have been isolated from each other since the end of the
J. J. ARMESTO, R. VERGARA and M. GARDNER (1999):
Genetic variation in Fitzroya cupressoides (alerce), athreatened South American conifer. Molecular Ecology
The high genetic distance between the artificially cul-
8: 975–987.
tivated TC population and the four wild populations
BEEBEE, T. J. C. and C. ROWE (2004): An introduction to
demonstrated that the seed source for this population is
molecular ecology. Oxford University Press, New York.
unlikely to have been any of the wild populations stud-
BROYLES, S. B. (1998): Postglacial migration and the loss
ied. According to the Flora of Yunnan (Kunming Insti-
of allozyme variation in northern populations of Ascle-
tute of Botany, 1986), there are still some wild popula-
pias exaltata (Asclepiadaceae). American Journal of
tions of T. cryptomerioides deep in the mountains, on the
Botany 85: 1091–1097.
border between China and Burma. The source of the TC
CHANG, J. L. (2005): The genetic diversity of Taiwania
population may be one of these unsampled populations
cryptomerioides Hayata. Master degree thesis. Taiwan:
or an extinct local wild population, since the local culti-
vation history of this species is very long and there are
CHUNG, Y. L. and N. H. CHANG (1990): Technical report for
many aged trees in the TC population. According to Far-
important tree seeds in Taiwan. Taiwan Forestry
jon (personal communication), the only endemic popula-
tions in mainland China are those near the border with
LARKE, G. M. and A. G. YOUNG (2000): Introduction:
genetics, demography and the conservation of fragment-
Burma, such as population GS in this study. However,
ed populations, pp. 1–6. In: Genetics, demography and
his opinion is not supported by the results of this study.
viability of fragmented populations, edited by A. G.
In addition, the populations located in the remote moun-
YOUNG and G. M. CLARKE, Cambridge University Press,
tains were inaccessible by road until the last two
decades. More work is needed to identify the source of
DOYLE, J. (1991): DNA protocols for plants CTAB total
the propagules from which LC, LS and GP were estab-
DNA isolation, pp. 283–293. In: Molecular techniques
lished. For most conifer species, mtDNA is inherited
in taxonomy, edited by G. M. HEWITT, A. JOHNSON and
maternally and is suitable for phylogeographical stud-
ESSELMAN, E. J., L. JIANQIANG, D. J. CRAWFORD, J. L. WIN-
endemic of China, using ISSR markers. Biochemical
DUS and A. D. WOLFE (1999): Clonal diversity in the rare
Genetics 44: 29–43. Calamagrostis porteri ssp. insperata (Poaceae): compar-
HOLSINGER, K. E. and P. O. LEWIS (2003): HICKORY: a
ative results for allozymes and random amplified poly-
package for analysis of population genetic data V1.0.
morphic DNA (RAPD) and intersimple sequence repeat
Department of Ecology and Evolutionary Biology, Uni-
(ISSR) markers. Molecular Ecology 8: 443–451.
versity of Connecticut, USA. Available at website
EXCOFFIER, L., P. E. SMOUSE and J. M. QUATTRO (1992):
http://darwin.eeb.uconn.edu/hickory/hickory.html
Analysis of molecular variance inferred from metric dis-
HU, Y. S., J. X. LIN, X. P. WANG and L. B. WEI (1995): The
tances among DNA haplotypes: application to human
biology and conservation of Taiwania cryptomerioides.
mitochondria DNA restriction sites. Genetics 131:
Chinese Biodiversity 3: 206–212.
HUANG, W. (1983): The vegetation of Taiwan. China Envi-
FARJON, A. (2001): World checklist and bibliography of
conifers, 2nd edn. England: The Royal Botanic Gardens
HUH, M. K. and H. W. HUH (2000): Genetic diversity and
population structure of Juniperus rigida (Cupressaceae)
FARJON, A. (2002): The discovery and protection of a Viet-
and J. coreana. Evolutionary Ecology 14: 87–98.
namese population of Taiwania cryptomerioides.
KUNMING INSTITUTE OF BOTANY (1986): Flora of Yunnan,
Species 38: 24.
FARJON, A. and S. O. GARCIA (2003). Cone and ovule
LEWONTIN, R. C. (1972): The apportionment of human
development in Cunninghamia and Taiwania (Cupres-
diversity. Evolutionary Biology 6: 381–398.
saceae sensu lato) and its significance for conifer evolu-
LI, F. G. and N. H. XIA (2005): Population structure and
tion. American Journal of Botany 90: 8–16.
genetic diversity of an endangered species, Glyp-
FENG, Z. Z., S. Z. YANG and D. M. WANG (1998): Rare trees
tostrobus pensilis (Cupressaceae). Botanical Bulletin of
in Yunnan Province. The China Esperanto Press, Bei-
Academia Sinica 46: 155–162.
LI, J. and Z. JIN (2007): Genetic variation and differentia-
FLORIN, R. (1963): The distribution of conifer and taxad
tion in Torreya jackii Chun, an endangered plant
genera in time space. Acta Horti Bergiana 20: 121–312.
endemic to China. Plant Science 172: 1048–1053.
FU, L. G. (1995): China Plant Red Data Book. Science
LI, L. C., S. SU and J. H. JIANG (2000): Karyotype analysis
of Chamaecparis obtusa var. formosana (Cupressaceae)
FU, L.G., Y. F. YU and A. FARJON (1999): Cupressaceae, pp.
and Taiwania cryptomerioides (Taxodiaceae). Journal of
62–65. In: Flora of China, edited by P. H. RAVEN and
Fudan University (Natural Science) 39: 569–571.
C. Y. WU, Science Press, and St. Louis: Missouri Botani-
LI, Y. Y., X. Y. CHEN, X. ZHANG, T. Y. WU, H. L. LU and
Y. W. CAI (2005): Genetic differences between wild and
GE, J. W. (1995): Preliminary study on Taiwania flou-
artificial populations of Metasequoia glyptostroboidessiana community in Maoba, Lichuan county, Hubei
Hu et Cheng (Taxodiaceae): Implications for species
Province, pp. 203–214. In: Vegetation of western Hubei
recovery. Conservation Biology 19: 224–231.
Province, edited by J. D. BAN, Huazhong University of
LIN, T. P., C. S. LU, Y. L. CHUNG and J. C. YANG (1993):
Allozyme variation in four populations of Taiwania
GE, S., D. Y. HONG, H. Q. WANG, Z. Y. LIU and C. M. ZHANG
in Taiwan. Silvae Genetica 42:
(1998): Population genetic structure and conservation
of an endangered conifer, Cathaya argyrophylla
LIU, L. H., J. H. ZHANG and Y. D. YU (1987): Studies on the
(Pinaceae). International Journal of Plant Sciences 159:
natural Taiwania flousiana forest and their communi-
ties in Yunnan province. Acta Phytoecologica Sinica 11:
GE, X. J., X. L. ZHOU, Z. C. LI, T. W. HSU, B. A. SCHAAL and
T. Y. CHIANG (2005): Low genetic diversity and signifi-
MELONI, M., D. PERINI, R. FILIGHEDDU and G. BINELLI
cant population structuring in the relict Amentotaxus
(2006): Genetic variation in five mediterranean popula-
argotaenia complex (Taxaceae) based on ISSR finger-
tions of Juniperus phoenicea as revealed by Inter-Sim-
printing. Journal of Plant Research 118: 415–422.
ple Sequence Repeat (ISSR) markers. Annals of Botany
GE, Y. Q, Y. X. QIU, B. Y. DING and C. X. FU (2003): An
97: 299–304.
ISSR analysis on population genetic diversity of the
MILLER, M. P. (1998): AMOVA-PREP 1.01: A program for
relict plant Ginkgo biloba. Biodiversity Science 11:
the preparation of AMOVA input files from dominant-
markers raw data. Computer software distributed by
GUPTA, M., Y. S. CHYI, J. ROMERO-SEVERSON and J. L.
OWEN (1994): Amplification of DNA markers from evolu-
NEI, M. (1973): Analysis of gene diversity in subdivided
tionarily diverse genomes using single primers of sim-
populations. Proceedings of the National Academy of
ple sequence repeats. Theoretical and Applied Genetics
Sciences of the United States of America. 70: 89: 998–1006.
HAMRICK, J. L., M. J. W. GODT and S. L. SHERMAN-BROYLE
NEWTON, A. C., T. ALLNUTT, A. C. M. GILLIES, A. LOWE and
(1992): Factors influencing levels of genetic diversity in
R. A. ENNOS (1999): Molecular phylogeography, intra-
woody plant species. New Forest 6: 95–124.
specific variation and the conservation of tree species.
HAMRICK, J. L. and M. J. W. GODT (1989): Allozyme diver-
Trends in Ecology and Evolution 14: 140–145.
sity in plant species, pp. 43–63. In: Plant population
NYBOM, H. (2004): Comparison of different nuclear DNA
genetics, breeding and genetic resources, edited by
markers for estimating intraspecific genetic diversity in
A. H. D. BROWN, M. T. CLEGG, A. L. KAHLER and B. S.
plants. Molecular Ecology 13: 1143–1155.
QIU, X. Q., S. Y. WU and K. H. LONG (1984): Investigation
HAO, B. Q., L. WANG, L. C. MU, L. YAO, R. ZHANG,
of Taiwania flousiana forests of Leigong mountain pre-
M. X. TANG and W. K. BAO (2006): A study of conserva-
serve in Guizhou Province. Acta Phytoecologica Sinica
tion genetics in Cupressus chengiana, an endangered
8: 264–278.
RENAU-MORATA, B. R., S. G. NEBAUER, E. SALES, J. ALLAIN-
WANG, X. P. and Y. K. LIU (1994): Theory and practice of
GUILLAUME, P. CALIGARI and J. SEGURA (2005): Genetic
biodiversity. China Environmental Science Press, Bei-
diversity and structure of natural and managed popula-
tions of Cedrus atlantica (Pinaceae) assessed using ran-
WOLFE, A. D. and A. LISTON (1998): Contributions of PCR-
dom amplified polymorphic DNA. American Journal of
based methods to plant systematics and evolutionary
Botany 92: 875–884.
biology, pp. 43–86. In: Plant molecular systematics II,edited by D. E. SOLTIS, P. S. SOLTIS and J. J. DOYLE,
SCHNEIDER, S., D. ROESSLI and L. EXCOFFIER (2000): Arle-
quin ver. 2.000: A software for population genetics data
WU, Z. Y. (1980): The vegetation of China. Science Press,
analysis, Genetics and Biometry Laboratory, University
YANG, Q. J., H. XU, Z. G. YAN, Y. LIU, K. G. ZHAO and
TANG, S. Q., W. J. DAI, M. S. LI, Y. ZHANG, Y. P. GENG,
L. Q. CHEN (2006): Natural resources and conservation
L. WANG and Y. ZHONG (2007): Genetic diversity of relic-
of Taiwania cryptomerioides in Hubei Province. Guihaia
tural and endangered plant Abies ziyuanensis26: 551–556.
(Pinaceae) revealed by AFLP and SSR markers. Geneti-
YANG, W. M. and S. Y. JIANG (1999): Investigation of relict
Taiwania flousiana in Fujian Province. Forest By-Prod- uct and Speciality in China 4: 55–57.
TSUMURA, Y., K. OHBA and S. H. STRAUSS (1996): Diversity
YEH, F. C., R. C. YANG and T. BOYLE (1999): POPGENE.
and inheritance of inter-simple sequence repeat poly-
Microsoft windows Based Freeware for Population
morphisms in Douglas-fir (Pseudotsuga menziesii) and
Genetic Analysis Release 1.31, University of Alberta,
sugi (Cryptomeria japonica). Theoretical and Applied
Genetics 92:
ZIETKIEWICZ, E., A. RAFALSKI and D. LABUDA (1994):
WALTHER, H. (1999): Die Tertiaerflora von Kleinsaubernitz
Genome fingerprinting by simple sequence repeat
bei Bautzen. Palaeontographica Abteilung B 249:
(SSR)-anchored polymerase chain reaction amplifica-
tion. Genomics 20: 176–183. Genetic Variation Within Two Sympatric Spotted Gum Eucalypts Exceeds Between Taxa Variation
By J. W. OCHIENG1),4),*), M. SHEPHERD1), P. R. BAVERSTOCK1), G. NIKLES2), D. J. LEE3) and R. J. HENRY1)
Abstract
implications for LD mapping, we investigated the level
Population substructure and hybridization, among
of molecular divergence between the two species at two
other factors, have the potential to cause erroneous
sympatric locations separated by 300 kilometres. Very
associations in linkage disequilibrium (LD) mapping.
few individuals of intermediate morphology were identi-
Two closely related spotted gum eucalypts, Corymbia
fied, despite the two species occurring only metres
variegata and C. henryi (Myrtaceae) occur in sympatry
apart. Analysis of genetic structure using 12 microsatel-
in the east coast of Australia and potentially interbreed.
lite loci showed that genetic differentiation between pop-
They are morphologically similar but are distinguished
ulations of the same species at different locations
as separate species based on capsule and foliage size. To
= 0.07 for both species; p = 0.0001) was significantly
determine whether they hybridize in nature and its
higher than that observed between species at each loca-tion (mean F
Bunyaville respectively; p = 0.0001; all Mann-Whitney
) Centre for Plant Conservation Genetics, Southern Cross Uni-
versity, P. O. Box 157 Lismore, NSW, 2480, Australia. U-test p ≤ 0.01). No species-specific alleles or significant
allele frequency differences were detected within a site,
) Department of Primary Industries and Fisheries, 80 Meiers
Road, Indooroopilly, Queensland, Australia.
suggesting recurr#ent local gene flow between the two
3) Department of Primary Industries and Fisheries, LB 16 Fraser
species. The lack of significant allele frequency differ-
Road, Gympie 4570, Queensland, Australia.
ences implies no population stratification along taxo-
4) Permanent address: Faculties of Agriculture & veterinary
nomic lines. This suggested that there is little concern
Medicine, University of Nairobi, P. O. Box 30197 Nairobi,
for cryptic hybridization when sampling from sites of
*) Communicating author: JOEL W. OCHIENG. Centre for Plant
Conservation Genetics, Southern Cross University, P. O. Box 57
Key words: panmixia, hybridization, gene flow, association
Lismore, NSW 2480 Australia. Tel: +61 2 6620 3961; Fax:
mapping, population structure, admixture, reproductive isola-
+61 2 6622 2080. E-mail: [email protected]
L’HYGIENE NEUROPSYCHIQUE Alimentation saine, sommeil régulier, temps de pause et pensées positives sont autant de mesures de bon sens pour être « bien dans sa tête ». S’il le fallait, la naturopathie apporte aussi des techniques naturelles fort utiles pour y arriver. La naturopathie holistique est la grande synthèse des méthodes naturelles de santé préventive et auto-curati
Changes in Depressive Symptoms and Social Functioning in theSequenced Treatment Alternatives to Relieve Depression StudyJohn W. Denninger, MD, PhD,* Adrienne O. van Nieuwenhuizen, MSc,* Stephen R. Wisniewski, PhD,ÞJames F. Luther, MSc,Þ Madhukar H. Trivedi, MD,þ A. John Rush, MD,§ Jackie K. Gollan, PhD,||Diego A. Pizzagalli, PhD,¶ and Maurizio Fava, MD*antidepressants, social functioning
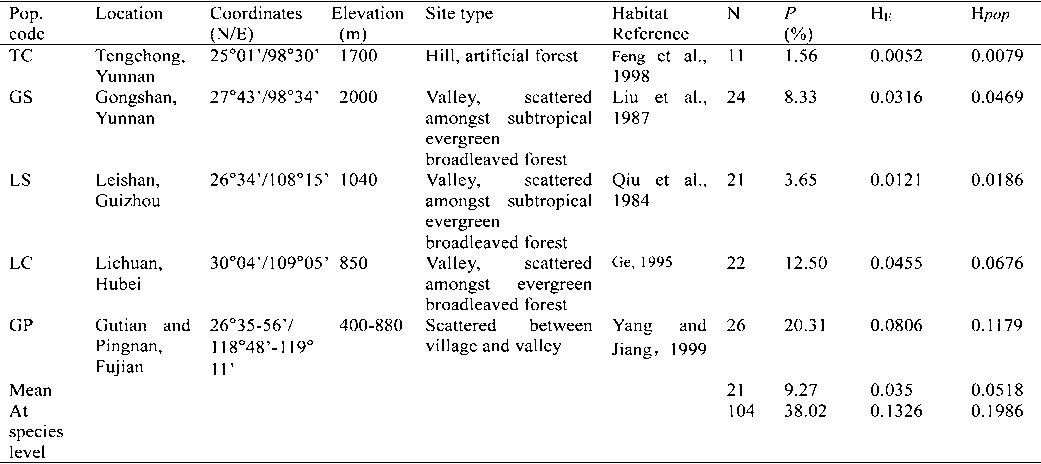
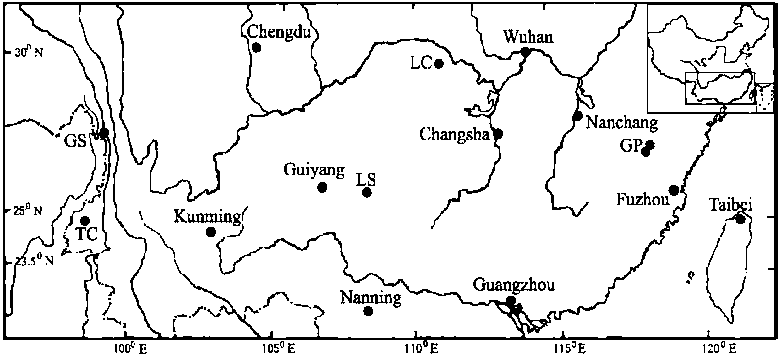 Table 1. – Genetic variability within populations of Taiwania cryptomerioides revealed by ISSR (N: sample size per population; P: percentage of polymorphic loci; H : Nei’s gene diversity; Hpop: Shannon indices).
Table 1. – Genetic variability within populations of Taiwania cryptomerioides revealed by ISSR (N: sample size per population; P: percentage of polymorphic loci; H : Nei’s gene diversity; Hpop: Shannon indices).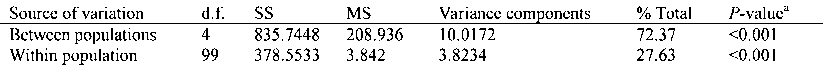
 individual. The resulting data matrix was analyzed
were 0.0518 at the population level (Hpop) and 0.1986
using POPGENE v. 1.31 (YEH et al., 1999) to estimate
at the species level (Hsp), respectively (Table 1). The cul-
genetic diversity parameters: the percentage of polymor-
tivated TC population exhibited the lowest genetic
phic loci (P) and expected heterozygosity (H ). At the
diversity. Among the four wild populations, population
species level, genetic diversity measures (H : total gene
GP had the greatest level of variability (P: 20.31%, H :
diversity; G : coefficient of gene differentiation) and the
0.0806, Hpop: 0.1179); population LS had the lowest
level of gene flow (Nm) were measured using Nei’s
level of variability (P: 3.65%, H : 0.0121, Hpop: 0.0186)
(1973) gene diversity statistics. Genetic diversity was
(Table 1). The average polymorphism of the four wild
also estimated using Shannon’s information statistic
= -Σpi log p where p is the fre-
The coefficient of genetic differentiation between pop-
ulations (G ) was 0.7269, estimated by partitioning the
two levels: the average diversity within the populations
total gene diversity. The Shannon’s diversity index
(Hpop), and the total diversity (Hsp). Then the propor-
analysis partitioned 73.92 % of the total variation
tion of diversity between the populations was estimated
between populations, in broad agreement with the
as: (Hsp-Hpop)/Hsp.
result of genetic differentiation analysis. The level of
In addition, an analysis of molecular variance
gene flow (Nm) was estimated to be 0.0939. The AMOVA
(AMOVA; EXCOFFIER et al., 1992) was used to estimate
analysis provided corroborating evidence for the genetic
variance components for ISSR phenotypes, partitioning
structure obtained from Nei’s genetic diversity statistics
the variation between populations and between individ-
and Shannon’s diversity estimate. There were highly
uals. AMOVA analysis was performed using the Arle-
significant (P < 0.001) genetic differences between the
quin 2.000 program (SCHNEIDER et al., 2000). To test the
five populations of T. cryptomerioides. Of the total mole-
presence of isolation by distance, a Mantel test between
cular variance, 72.37 % was attributable to between-pop-
the pairwise genetic distance and geographic distance
ulation diversity and the rest (27.63 %) to differences
from each population pair was applied using Mantel’s
between individuals within populations (Table 2). A sim-
test in the software TFPGA (MILLER, 1997) (1000 per-
ilar result was obtained from the Hickory calculation: θ B
mutations). Pairwise genetic distances (Φ ) were got
was 0.668 for the f free model, which had the smallest
from AMOVA analysis, which are analogous to tradition-
Genetic distances (Φ ) between populations of
An alternative Bayesian method, allowing direct esti-
T. cryptomerioides were calculated by Arlequin 2.000
(SCHNEIDER et al., 2000) (Table 3). The highest distance
ous knowledge of F , was employed to estimate θ B
value was 0.9011 between TC and LS, and the lowest
(analogous to F ) using Hickory 1.0 (HOLSINGER and
was 0.5037 between LS and GP. Geographically TC and
LEWIS, 2003). Using the default sampling parameters
GS are the closest pair of sites (269 km), however, the
(burn-in = 5 000, sample = 25 000, thin = 5), θ B was cal-
genetic distance between them, 0.7608, was not the low-
culated under four different models. The first was a full
est of all the pairs. The result of a Mantel test with 1000
model, in which both θ B and f (i.e., F , the inbreeding
permutations revealed that the genetic divergence of
coefficient) were estimated. Two alternative models
populations was not significantly correlated with geo-
assumed either θ B or f equal to zero. Finally, because
graphical distance (Mantel test, r = -0.3124, P = 0.2170).
estimates of f based on dominant markers are usuallystrongly biased (especially at low sample sizes, i.e.,n < 10), HICKORY was used to construct a model in
Discussion
individual. The resulting data matrix was analyzed
were 0.0518 at the population level (Hpop) and 0.1986
using POPGENE v. 1.31 (YEH et al., 1999) to estimate
at the species level (Hsp), respectively (Table 1). The cul-
genetic diversity parameters: the percentage of polymor-
tivated TC population exhibited the lowest genetic
phic loci (P) and expected heterozygosity (H ). At the
diversity. Among the four wild populations, population
species level, genetic diversity measures (H : total gene
GP had the greatest level of variability (P: 20.31%, H :
diversity; G : coefficient of gene differentiation) and the
0.0806, Hpop: 0.1179); population LS had the lowest
level of gene flow (Nm) were measured using Nei’s
level of variability (P: 3.65%, H : 0.0121, Hpop: 0.0186)
(1973) gene diversity statistics. Genetic diversity was
(Table 1). The average polymorphism of the four wild
also estimated using Shannon’s information statistic
= -Σpi log p where p is the fre-
The coefficient of genetic differentiation between pop-
ulations (G ) was 0.7269, estimated by partitioning the
two levels: the average diversity within the populations
total gene diversity. The Shannon’s diversity index
(Hpop), and the total diversity (Hsp). Then the propor-
analysis partitioned 73.92 % of the total variation
tion of diversity between the populations was estimated
between populations, in broad agreement with the
as: (Hsp-Hpop)/Hsp.
result of genetic differentiation analysis. The level of
In addition, an analysis of molecular variance
gene flow (Nm) was estimated to be 0.0939. The AMOVA
(AMOVA; EXCOFFIER et al., 1992) was used to estimate
analysis provided corroborating evidence for the genetic
variance components for ISSR phenotypes, partitioning
structure obtained from Nei’s genetic diversity statistics
the variation between populations and between individ-
and Shannon’s diversity estimate. There were highly
uals. AMOVA analysis was performed using the Arle-
significant (P < 0.001) genetic differences between the
quin 2.000 program (SCHNEIDER et al., 2000). To test the
five populations of T. cryptomerioides. Of the total mole-
presence of isolation by distance, a Mantel test between
cular variance, 72.37 % was attributable to between-pop-
the pairwise genetic distance and geographic distance
ulation diversity and the rest (27.63 %) to differences
from each population pair was applied using Mantel’s
between individuals within populations (Table 2). A sim-
test in the software TFPGA (MILLER, 1997) (1000 per-
ilar result was obtained from the Hickory calculation: θ B
mutations). Pairwise genetic distances (Φ ) were got
was 0.668 for the f free model, which had the smallest
from AMOVA analysis, which are analogous to tradition-
Genetic distances (Φ ) between populations of
An alternative Bayesian method, allowing direct esti-
T. cryptomerioides were calculated by Arlequin 2.000
(SCHNEIDER et al., 2000) (Table 3). The highest distance
ous knowledge of F , was employed to estimate θ B
value was 0.9011 between TC and LS, and the lowest
(analogous to F ) using Hickory 1.0 (HOLSINGER and
was 0.5037 between LS and GP. Geographically TC and
LEWIS, 2003). Using the default sampling parameters
GS are the closest pair of sites (269 km), however, the
(burn-in = 5 000, sample = 25 000, thin = 5), θ B was cal-
genetic distance between them, 0.7608, was not the low-
culated under four different models. The first was a full
est of all the pairs. The result of a Mantel test with 1000
model, in which both θ B and f (i.e., F , the inbreeding
permutations revealed that the genetic divergence of
coefficient) were estimated. Two alternative models
populations was not significantly correlated with geo-
assumed either θ B or f equal to zero. Finally, because
graphical distance (Mantel test, r = -0.3124, P = 0.2170).
estimates of f based on dominant markers are usuallystrongly biased (especially at low sample sizes, i.e.,n < 10), HICKORY was used to construct a model in
Discussion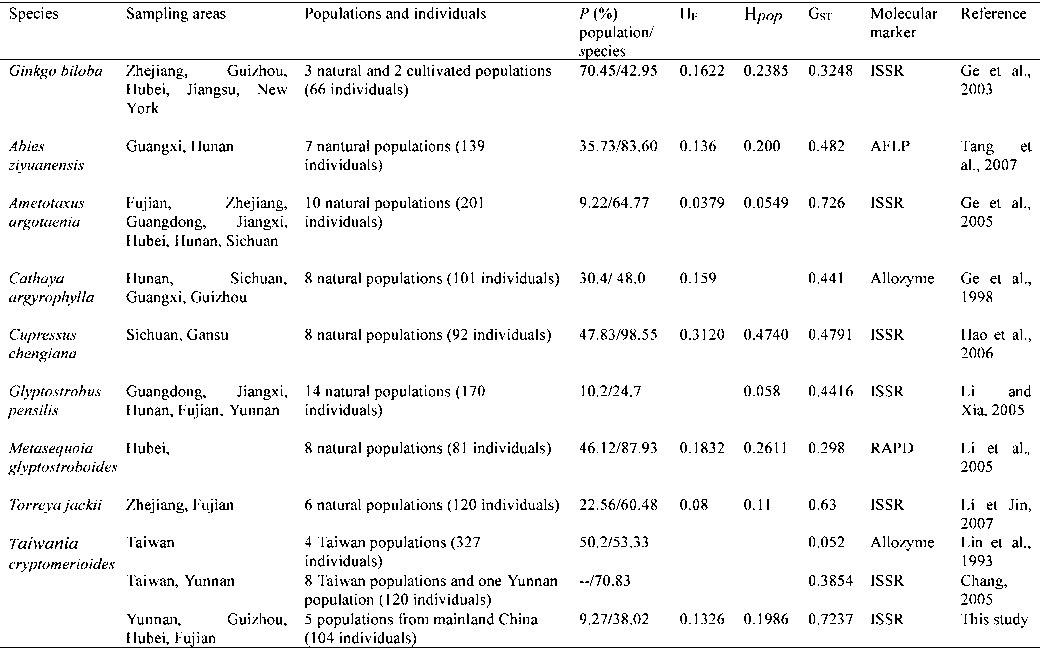 on a vast amount of data relating to natural plant popu-
pensilis (LI and XIA, 2005), Metasequoia glyptostroboides
lations (HAMRICK and GODT, 1989; NYBOM, 2004). Most
(LI et al., 2005), Cupressus chengiana (HAO et al., 2006),
conifers have high levels of genetic diversity and low
Torreya jackki (LI and JIN, 2007) and Abies ziyuanensis
levels of differentiation between populations (HAMRICK
(TANG et al., 2007) (Table 4). Since these species are
et al., 1992). Examples include Fitzroya cupressoides
characterized by their relict status, biogeographic histo-
(Hpop: 0.547, Φ : 0.1438; ALLNUTT et al., 1999), Junipe-
ry may play an important role in determining their
rus rigida, J. coreana (H : 0.224 and 0.199; G : 0.173
genetic diversity. T. cryptomerioides is one of the repre-
and 0.118, respectively; HUH and HUH, 2000), Cedrus
sentatives of relict gymnosperms from the Tertiary peri-
atlantica (Nei’s gene diversity within population, H :
od that are found in China. Fossil evidence indicates
0.194; F : 0.148; θ B: 0.178; RENAU-MORATA et al., 2005)
that the ancestor of T. cryptomerioides was much more
and Juniperus phoenicea (H : 0.148, H : 0.130, G :
widespread during the Tertiary than its current distrib-
0.12; MELONI et al., 2006). Similar genetic diversity and
ution suggests (WALTHER, 1999). Accordingly, a reason-
genetic differentiation have also been reported for the
able hypothesis is that the modern range of T. cryptome-
Taiwanese populations of T. cryptomerioides (LIN et al.,
rioides was the result of population fragmentation and
1993; CHANG, 2005). In this study, however, ISSR poly-
contraction after the Quaternary glacial cycles. The
morphisms revealed a low level of genetic diversity in
remnant populations contained only part of the genetic
the mainland populations of T. cryptomerioides, with an
variation present before the range reduction. Genetic
average of only 11.19% of ISSR bands being polymorphic
drift and inbreeding may have further decreased the
in the four wild populations studied. In addition, strong
genetic diversity and increased genetic differentiation in
population differentiation was demonstrated based on
the small, isolated populations of T. cryptomerioides in
Nei’s gene diversity, Shannon’s information statistic,
AMOVA and Bayesian estimates. About 70 % of the total
In addition to historical influences, the present small
genetic variation was partitioned between the popula-
population size of T. cryptomerioides may be important.
on a vast amount of data relating to natural plant popu-
pensilis (LI and XIA, 2005), Metasequoia glyptostroboides
lations (HAMRICK and GODT, 1989; NYBOM, 2004). Most
(LI et al., 2005), Cupressus chengiana (HAO et al., 2006),
conifers have high levels of genetic diversity and low
Torreya jackki (LI and JIN, 2007) and Abies ziyuanensis
levels of differentiation between populations (HAMRICK
(TANG et al., 2007) (Table 4). Since these species are
et al., 1992). Examples include Fitzroya cupressoides
characterized by their relict status, biogeographic histo-
(Hpop: 0.547, Φ : 0.1438; ALLNUTT et al., 1999), Junipe-
ry may play an important role in determining their
rus rigida, J. coreana (H : 0.224 and 0.199; G : 0.173
genetic diversity. T. cryptomerioides is one of the repre-
and 0.118, respectively; HUH and HUH, 2000), Cedrus
sentatives of relict gymnosperms from the Tertiary peri-
atlantica (Nei’s gene diversity within population, H :
od that are found in China. Fossil evidence indicates
0.194; F : 0.148; θ B: 0.178; RENAU-MORATA et al., 2005)
that the ancestor of T. cryptomerioides was much more
and Juniperus phoenicea (H : 0.148, H : 0.130, G :
widespread during the Tertiary than its current distrib-
0.12; MELONI et al., 2006). Similar genetic diversity and
ution suggests (WALTHER, 1999). Accordingly, a reason-
genetic differentiation have also been reported for the
able hypothesis is that the modern range of T. cryptome-
Taiwanese populations of T. cryptomerioides (LIN et al.,
rioides was the result of population fragmentation and
1993; CHANG, 2005). In this study, however, ISSR poly-
contraction after the Quaternary glacial cycles. The
morphisms revealed a low level of genetic diversity in
remnant populations contained only part of the genetic
the mainland populations of T. cryptomerioides, with an
variation present before the range reduction. Genetic
average of only 11.19% of ISSR bands being polymorphic
drift and inbreeding may have further decreased the
in the four wild populations studied. In addition, strong
genetic diversity and increased genetic differentiation in
population differentiation was demonstrated based on
the small, isolated populations of T. cryptomerioides in
Nei’s gene diversity, Shannon’s information statistic,
AMOVA and Bayesian estimates. About 70 % of the total
In addition to historical influences, the present small
genetic variation was partitioned between the popula-
population size of T. cryptomerioides may be important.