Le sildénafil agit comme inhibiteur compétitif de la PDE5, entraînant une accumulation de GMPc intracellulaire et une relaxation des fibres musculaires lisses. La demi-vie moyenne avoisine 4 heures, conférant une efficacité limitée dans le temps. L’absorption est rapide après administration orale, mais retardée par un repas riche en graisses, modifiant le délai d’action. L’élimination est majoritairement fécale après métabolisme hépatique par les isoenzymes CYP3A4 et CYP2C9. Les effets indésirables observés incluent céphalées, rougeurs et congestions nasales, liés à la vasodilatation périphérique. Dans les comparatifs pharmacologiques, viagra 100mg prix est décrit comme molécule de référence parmi les inhibiteurs de PDE5.
Created using pdfonline.com , a free pdf creation service
Quantum Chemical Design of Hydroxyurea Derivatives For the Treatment of Sickle Cell Anemia I. Introduction
Sickle cell anemia is an inherited disorder in which red blood cells become stiff
and sickle-shaped. This condition is caused by defective hemoglobin that clusters
together, forming long, rod-like structures.1 The abnormal red blood cells cannot freely
move through small blood vessels and thus cause blockages that deprive organs and
tissues of oxygen.2,3 A study published in 2003 established that the use of hydroxyurea
therapy decreases mortality among sickle cell patients by forty percent and significantly
reduces pain and acute chest crises.2 Hydroxyurea produces an increase of fetal
hemoglobin, which prevents the polymerization of sickle hemoglobin.1,4 It is also a
source of nitric oxide (NO), a messenger molecule needed to maintain normal blood flow
and pressure.4-8 Hydroxyurea reacts with hemoglobin by first forming a nitroxide
radical.6 It then undergoes a series of reactions to produce the nitric oxide needed to
increase fetal hemoglobin.4,6,7 Although the production of NO can proceed through
various pathways, the process always requires the removal of the hydrogen atom from the
This study concerns the formation of the nitroxide radical, which is the rate-
limiting reaction in the process by which hydroxyurea treats sickle cell disease.5 Huang,
Kim-Shapiro, and King have recently shown by experiment that derivatives of the
hydroxyurea molecule can form NO-producing radicals more quickly than hydroxyurea
can produce nitroxide.7 In this work, the ability of different hydroxyurea derivates to
generate NO-producing radicals is explored computationally. The molecular energies of
hydroxyurea derivatives and their corresponding radicals were computed using a variety
of quantum chemical methods, including Hartree-Fock theory, density functional theory,
and correlated wavefunction (or ab initio) methods, such as many-body perturbation
theories and coupled-cluster methods. Specifically, energy difference between
hydroxyurea and the nitroxide radical was compared with the energy difference between
each hydroxyurea derivative and its radical. As explained in this report with
thermodynamic and kinetic arguments, the smaller energy differences favor the formation
of the radical. A table “ranks” the derivatives according to their energy differences and
compares the results with experimentally determined rate constants when available. The
results of the study show that all nineteen hydroxyurea derivatives form NO-producing
radicals faster than hydroxyurea. One of these derivatives, Zileuton, approved by the
Food and Drug Administration for the treatment of asthma, has been shown to increase
fetal hemoglobin in sickle cell patients.9
II. Background
Hydroxyurea produces nitric oxide (NO) through a three-electron oxidation.6
This requires a single-electron oxidation of hydroxyurea to first produce the nitroxide
radical, which may disproportionate to form NO (Figure 1). The radical may also
undergo another single-electron oxidation, forming C-nitrosoformamide before
Figure 1: Hydroxyurea produces the nitroxide radical, which then disproportionates to form NO.
Hydroxyurea forms the nitroxide radical by interacting with hemoglobin.1 First, it
oxidizes oxyhemoglobin (Fe+2-O2) to yield methemoglobin (Fe+3) and the nitroxide
radical. Hydroxyurea then reacts with methemoglobin, forming a low-spin
methemoglobin-hydroxyurea complex.5,6 This complex produces deoxyhemoglobin and
another nitroxide radical that decomposes to nitric oxide as previously described (Figure
2). Hydroxyurea can increase fetal hemoglobin levels by using the nitric oxide produced
through a guanylate cyclase pathway as well as through many other pathways.4,7 The
reactions described occur only at moderate rates and require a large excess of
hydroxyurea,7 suggesting that other molecules similar to hydroxyurea may be more
Figure 2: Hydroxyurea forms the nitroxide radical by oxidizing oxyhemoglobin to methemoglobin. It also reacts with the methemoglobin to form a methemoglobin complex, which then produces deoxyhemoglobin and another nitroxide radical. III. Methodology
In the human body, the conversion of hydroxyurea to the nitroxide radical is
assisted by the reaction of hemoglobin, either from oxyhemoglobin to methemoglobin or
from methemoglobin to deoxyhemoglobin. Alone, the removal of the hydrogen atom
from hydroxyurea to form the nitroxide radical requires such a considerable amount of
energy that virtually no nitroxide would form, but because the transformation of
hemoglobin is thermodynamically favorable, the probability of hydroxyurea to produce
the nitroxide radical increases within the body. Additionally, the removal of nitroxide by
its decomposition to nitric oxide assists nitroxide production because, by LeChatlier’s
principle, the reaction compensates for the loss of product by creating more product.
However, a large excess of hydroxyurea is necessary for the reaction to occur because the
equilibrium constant still significantly favors hydroxyurea.
Because the removal of a hydrogen atom from a hydroxyurea derivative does not
cause an appreciable change in pressure or volume of the system, the change in enthalpy
is equal to the energy difference. Furthermore, since the entropy for the reaction also
remains essentially constant, the free energy change is equivalent to the change in
electronic energy (Figure 3). The electronic energy differences between the hydroxyurea
derivatives and their radicals alone indicate the favorability of their equilibrium constants
for radical formation because the ratio of the equilibrium constant for the conversion of
hydroxyurea to nitroxide to the equilibrium constant for the conversion of a hydroxyurea
derivative to its radical is independent of any hemoglobin reactions. Hemoglobin has the
same thermodynamic role in both reactions, and thus, its concentration cancels in the
ratio of the equilibrium constants for the two reactions, as shown in Figure 4. This means
that comparing electronic energy differences between derivatives and their radicals is an
accurate method for ordering the molecules based on their thermodynamic formation of
Consider the relations ΔH = ΔE + Δ(PV) and ΔG = ΔH - T ΔS. Figure 3: If there are no changes in pressure, volume, or entropy, the free energy change may be equated to the change in energy between hydroxyurea or any of its derivatives and their corresponding radical.
K2 = [methemoglobin] [peroxide] / [oxyhemoglobin]
K3 = [radical] / [hydroxyurea derivative]
Figure 4: Let Kx represent the equilibrium constant for the conversion of hydroxyurea to nitroxide, and let Ky represent the equilibrium constant for the conversion of a hydroxyurea derivative to its radical. Dividing Kx by Ky will cause the equilibrium constant for the hemoglobin reactions cancel out. Therefore, the reactions of hydroxyurea and its derivatives may be compared with one another without considering the hemoglobin reactions.
Without hemoglobin, the potential energy surface of the process of removing
hydrogen from hydroxyurea increases monotonically with the distance of the hydrogen
atom from the remaining hydroxyurea molecule. In the presence of hemoglobin,
therefore, the removal of hydrogen would be expected to have an energy barrier
corresponding to the hydroxyurea-hemoglobin transition state. The reaction rates of the
different hydroxyurea derivatives will consequently depend on the height of the
transition-state barrier. To compare these rates, it can be assumed that changes in the
energy difference between hydroxyurea derivatives and their radicals translate to similar
changes in the barrier height. Although equilibrium constants may be compared without
consideration of the interactions between the hydroxyurea derivative and hemoglobin,
reaction rates may be influenced by such interactions between the hydroxyurea derivative
and hemoglobin, such as steric effects or kinetic enhancements. Comparing only
hydroxyurea derivatives and their radicals allows us to determine an estimate of the
relative reaction rates from well-defined electron structure calculations without the
treatment of hemoglobin in the transition state.
Various methods were employed to calculate the electronic energies of the
hydroxyurea derivatives and their radicals. In addition to Hartree-Fock, correlated
wavefunction methods were used, including second- and fourth-order perturbation
theories (MP2 and MP4), coupled-cluster singles-doubles (CCSD), and coupled cluster
singles-doubles with a perturbative triples correction (CCSD(T)). Three different
correlation-exchange functions (PBEPBE, G96PBE, and B3LYP) were used within
density functional theory. For each method, calculations were done in three Pople basis
sets (6-31G, 6-31G*, and 6-31G**) using the quantum chemistry code Gaussian.10
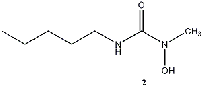
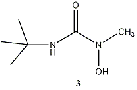
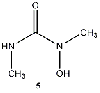
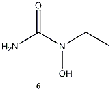
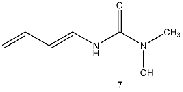
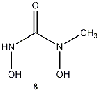
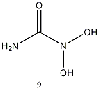
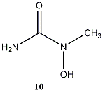
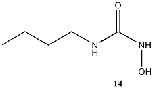
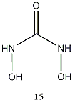
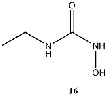
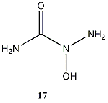
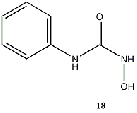
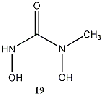
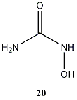
Equilibrium geometries were found for all molecules at the Hartree-Fock level of theory. Hydroxyurea (20) and nineteen of its derivatives were tested in this study, including Zileuton (1), an asthma medication that has been shown to increase fetal hemoglobin in sickle cell patients. Derivatives 8 and 19 have the same structure, but the radical of derivative 8 forms on the CH3 side, while the radical of derivative 19 forms on the opposite side. IV. Calculations and Results
Hydroxyurea and nineteen derivatives were tested in this study (Table I),
including some of those proposed and chemically tested by S. B. King.5 Every derivative
was investigated using the previously described methods. Table II provides a comparison
of the energy difference between hydroxyurea and nitroxide with the energy difference
between hydroxyurea derivative 10 and its radical. The results of using three correlated
wavefunction methods are given as well as the calculations using three density functional
methods. Specifically, second-order many-body perturbation theory (MP2), coupled
cluster singles doubles (CCSD), and coupled cluster singles doubles with a pertubative
triples correction (CCSD(T)) are the correlated methods shown, and PBEPBE, G96PBE,
and B3LYP are the density functional methods shown. The Hartree-Fock (HF) method,
which does not include correlation energy, is also displayed. This method predicts that
the modification of hydroxyurea will lower the energy difference between the molecule
and the radical by 29.5 kJ/mol, whereas the correlated wavefunction methods predict a
larger decrease in the energy difference. The MP2, CCSD, and CCSD(T) methods yield
an energy reduction by 20.6 kJ/mol, 64.4 kJ/mol, and 106.8 kJ/mol respectively. The
density function methods, PBEPBE, G96PBE, and B3LYP, show a decrease in the
energy differences by 51.7 kJ/mol, 51.4 kJ/mol, and 48.8 kJ/mol respectively. The
energy methods with electron correlation included predict a much larger difference
between the molecules and their radicals. Correlation energy accounts for more than fifty
percent of the energy difference between molecules and radicals for some of the
hydroxyurea derivatives. The density function theory calculations tend to indicate an
even larger energy difference than the correlated wavefunction methods. The table
shows these differences among calculations performed in a 6-31G** basis set.
Calculations done in the 6-31G and 6-31G* basis sets produced similar trends.
Table III provides a comparison of the energy differences between the molecule
and radical found using the B3LYP density functional method in basis sets 6-31G, 6-
31G*, and 6-31G**. All the molecules, arranged in order of increasing energy
difference, have smaller energy differences than hydroxyurea (molecule 20). Derivatives
with a group substituted for one or both of the hydrogen atoms on the side opposite the
hydroxide group showed the smallest improvement over hydroxyurea in the energy
difference between molecule and their derivative (12-16, 18). Of these molecules, the
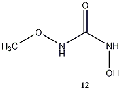
methyl ether derivative (12) produced the least energy difference, while the derivative
with a butyl group (14) and well as one with an ethyl group (16) also produced some of
the smaller energy differences. The least improvement was observed from the
substitution of a phenyl group (18).
Substitutions on the hydroxide side of the molecule tended to produce smaller and
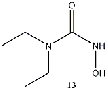
thus more favorable energy differences. The addition of an ethyl group, a hydroxide
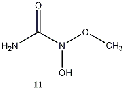
group (9), methyl group (10), or a methyl ether group (11) greatly improved the energy
difference. Within this class, an amine substitution (17) produced the least improvement
in energy difference, while the hydroxyurea derivative Zileuton (1) yielded the most
significant improvement over hydroxyurea in the study. Although Zileuton’s energy
difference of 1608.8 kJ/mol might not seem much more favorable than the 1670.0 kJ/mol
energy difference of hydroxyurea, it is important because small changes in energy have
large and significant effects on equilibrium and kinetics.
The derivatives with substitutions on both nitrogen atoms (2-5, 7, 8) generally
produce even smaller energy differences between molecule and radical. Each has a
methyl group attached to the hydroxide side. The substitution of a phenyl group (2)
produces the most favorable results, although the addition of a butyl (4) or methyl group
(5) also yields a very small molecule–radical energy difference. A branched alkane
substitution (3) slightly improved the difference from the butyl group (4), while an
unsaturated chain (7) yielded a greater energy difference. Although longer alkane chains
generally produce small molecule-radical energy differences, the steric effect from
additional chains may be unfavorable for the transition state.
The experimental rate constants available for five molecules tested in this study
shown in Table III (10, 13, 14, 18, 20) generally agree with these calculations. The rate
constant for the formation of nitroxide from hydroxyurea (7.54 x 10-4 min-1) is slower
than any of the derivatives, which agrees with the results of this study. Derivative 10
generates its radical about two hundred times faster than hydroxyurea. This experimental
result highlights how small energy differences, in this case a 48.8 kJ/mol difference in
molecule-radical energy gaps between hydroxyurea and derivative 10, have significant
effects on reaction rates. The experimental rate constants also agree with the calculations
for derivatives 10, 13, and 14, although derivative 18 has a much more favorable rate
constant than the one expected from its energy calculations. Its rate is similar to
derivative 10 while its computed energy difference is larger by approximately 24.2
kJ/mol. This discrepancy may have arisen because the phenyl group may have
kinetically enhanced the transition state through a stabilization. This effect cannot be
predicted without examination of the methemoglobin-hydroxyurea complex. The molecule-radical energy difference of hydroxyurea is greater than that of Derivative 10, suggesting that Derivative 10 may form a radical more quickly than hydroxyurea.
Energy Differences (kJ/mol) between Hydroxyurea
Energy Differences (kJ/mol) between Derivative
Table III Hydroxyurea derivatives are ranked in order of increasing molecule-radical energy differences in three basis sets. Any known experimental rate constants are also provided.
Energy Differences (kJ/mol) between Hydroxyurea Derivatives and their Radicals
V. Conclusions
This study has investigated the effects of modifying hydroxyurea to increase the
generation of nitric oxide-producing radicals for the treatment of sickle cell anemia.
Electronic energies were calculated for derivatives of hydroxyurea and their radicals, and
the energy differences between the molecules and the radicals have been used to compare
their thermodynamic and kinetic favorability for radical formation by hydrogen
abstraction. Every derivative tested had a smaller and thus more favorable energy
difference than hydroxyurea. Alone, this energy difference cannot exactly predict the
reaction rate of radical formation because the transition state involves both hydroxyurea
and hemoglobin. The hemoglobin-hydroxyurea interactions may produce kinetic
enhancements. However, to a first-order of approximation, the energy differences
between each molecule and its radical offer an estimate of the molecule-radical
conversion rate. Experimental rate constants confirm that some derivatives of
hydroxyurea form radicals more quickly than hydroxyurea. A computational
investigation has significant advantages because many molecules can be tested without a
time-consuming and possibly expensive laboratory synthesis.
The results of this study indicate that correlation energy significantly affects the
magnitude of electronic energy calculations. The energy differences between each
molecule and its radical are significantly larger with electron correlation. It can be
concluded, therefore, that correlation energy is crucial to accurately compare energy
differences between molecules. Both the basis set size and the correlation method affects
the calculated energy differences, but these changes are small compared to those from the
The results of this study suggest that hydroxyurea derivatives may form radicals
at a faster rate than hydroxyurea. In particular, the asthma medication Zileuton seems to
have the fastest reaction rate of any derivative tested. A recent study also indicates that
this drug increases fetal hemoglobin in sickle cell patients. The derivatives of
hydroxyurea may also improve treatment of sickle cell anemia by producing fewer
undesirable side effects. To fully determine the affects of substituting a hydroxyurea
derivative for hydroxyurea to treat sickle cell disease, the activity of the molecule during
the reactions following radical formation must also be investigated. Computation of
hydroxyurea derivatives and their radicals by quantum chemistry provides a unique
approach to considering modifications of hydroxyurea for improved treatment of sickle-
_________________________________________
References
(1) M. W. Rupon, S. R. Domingo, S. V. Smith, et al, Biophysical Chemistry, 84, 1-8, 2000.
(2) L. De Franceschi and P. G. B. Rossi, Haematologica, 89, 348-353 (2004).
(3) S. K. Ballas and E. D. Smith, Blood, 79, 2154-2158 (1992).
(4) V. P. Cokic, R. D. Smith, B. B. Beleslin-Cokic, et al, J. Clin. Invest. 111, 231-235
(5) J. Huang, Z. Zou, D. B. Kim-Shapiro, S. K. Ballas, and S. B. King, J. Med. Chem. 46,
(6) S. B. King, Current Medical Chemistry, 10, 437 (2003).
(7) J. Huang, D. B. Kim-Shapiro, and S. B. King, J. Med. Chem. 47, 3496 (2004).
(8) C. Halsey and I. A. G. Roberts, Br. J. Haematology, 120, 177-182, 2003.
(9) J. Haynes Jr, B. S. Baliga, B. Obiako, S. Ofori-Acquah, and B. Pace, Blood, 103, 3945
(10) M. J. Frisch, G. W. Trucks, H.B. Schlegel et al., Gaussian 98 (Gaussian, Inc.,
MANDATORY GEAR LIST The following is a list of equipment that you should have, at a minimum, for the duration of the event. Mandatory Gear will be checked at Registration before the event starts Spot checks will take place before participants start each day For your own safety and in the interests of fair play, make sure you have all Mandatory Gear before you leave the Start or O
Alpine Medical Clinic 211 Bear St, Unit 201A, Banff, Alberta T: (403)762-3155 F: (403)762-5797 www.banffmedical.com CARE OF YOUR HEAD INJURY/CONCUSSION General Information A mild head injury also known as a concussion is a brain injury that cannot be seen on routine x-rays, CT scans or MRIs. It is caused by striking the head against an object or by a blow to the head. It affects the way a
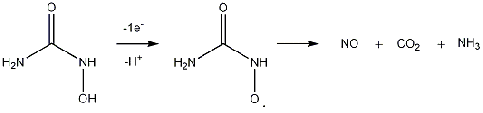 Figure 1: Hydroxyurea produces the nitroxide radical, which then disproportionates to form NO.
Figure 1: Hydroxyurea produces the nitroxide radical, which then disproportionates to form NO.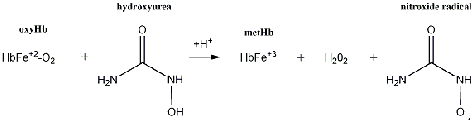
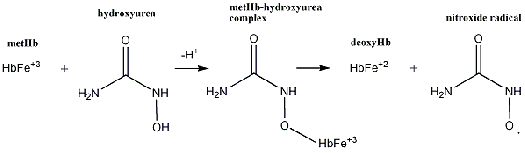 Figure 2: Hydroxyurea forms the nitroxide radical by oxidizing oxyhemoglobin to methemoglobin.
Figure 2: Hydroxyurea forms the nitroxide radical by oxidizing oxyhemoglobin to methemoglobin. 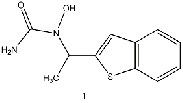
 and hemoglobin, such as steric effects or kinetic enhancements. Comparing only
hydroxyurea derivatives and their radicals allows us to determine an estimate of the
relative reaction rates from well-defined electron structure calculations without the
treatment of hemoglobin in the transition state.
and hemoglobin, such as steric effects or kinetic enhancements. Comparing only
hydroxyurea derivatives and their radicals allows us to determine an estimate of the
relative reaction rates from well-defined electron structure calculations without the
treatment of hemoglobin in the transition state.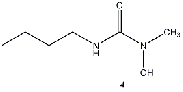
 IV. Calculations and Results
IV. Calculations and Results